| Le basi della genetica |
|---|
|
| | Aplogruppi DNA-Y |
|---|
|
| | Aplogruppi del mtDNA |
|---|
|
| | Storie genetiche regionali |
|---|
|
| | Cronologia genetica |
|---|
|
| | DNA autosomica |
|---|
|
| | Altre pagine sul DNA |
|---|
|
|
| |
Autore: Maciamo Hay. Traduzione italiana da Enea Sagittario.
Distribuzione geografica
Distribuzione europea dellaplogruppo R1b

Distribuzione dellaplogruppo R1b nel Vecchio Mondo
 - Eupedia")
Filogenesi di R1b
Se non avete dimestichezza con la genealogia genetica per favore controllate la nostra Introduzione alla filogenetica per meglio comprendere come leggere un albero filogenetico.
 - Eupedia")
Clicca sugli alberi sottostanti per ingrandire
 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")
Origini e storia
Cacciatori di mammut paleolitici
Laplogruppo R* ebbe origine in Asia settentrionale poco prima dellultimo massimo glaciale (26.500-19.000 anni fa). È stato identificato nei resti di un ragazzo vissuto circa 24.000 anni fa nella regione dei monti Altaj, in Siberia centromeridionale (Raghavan et al. 2013). Questo individuo apparteneva a una tribù di cacciatori di mammut che, durante il Paleolitico, conducevano una vita nomade tra la Siberia e parte dellEuropa. Secondo lanalisi autosomica, questa popolazione paleolitica sembra abbia contributo soprattutto a determinare lattuale composizione genetica degli Europei moderni e degli abitanti dellAsia meridionale, le due regioni dove anche oggi laplogruppo R è più diffuso (R1b in Europa occidentale, R1a in Europa orientale e in Asia centrale e meridionale, R2 in Asia meridionale).
Le forme più antiche di R1b (M343, P25, L389) si trovano oggi disperse con basse frequenze in unarea compresa tra lEuropa occidentale e lIndia, una regione molto vasta dove, durante lera glaciale, avrebbero vagabondato le tribù di cacciatori nomadi R1b. Sembra che tutti e tre i rami principali di R1b1 (R1b1a, R1b1b, R1b1c) abbiano avuto origine in Medioriente. Il ramo meridionale, R1b1c (V88 o R1b1a2), si trova oggi soprattutto nel Levante e in Africa. Il ramo settentrionale, R1b1a (P297 o R1b1a1a), sarebbe apparso da qualche parte tra il Caucaso, lAnatolia orientale e la Mesopotamia settentrionale; da qui si sarebbe poi diffuso proprio nel Caucaso per poi colonizzare lEuropa e lAsia centrale. Il terzo ramo, R1b1b (M335 o R1b2), fino ad oggi è stato trovato solo in Anatolia.
Mandriani neolitici
È stato ipotizzato che, circa 10.500 anni fa, i bovini siano stati addomesticati per la prima volta nella storia dalle tribù R1b della Mesopotamia settentrionale, probabilmente insieme ai loro vicini J2. Queste genti R1b discendevano dai più antichi cacciatori di mammut che, quando questi animali si estinsero, cominciarono a cacciare soprattutto bisonti e uri. Nella Mezzaluna fertile, anche grazie allincremento demografico cominciato con il neolitico (12.000 anni fa), fu necessario sostituire alla caccia indiscriminata una caccia più selettiva. Linfluenza dellattività umana sulla vita degli uri, dei maiali selvatici e delle capre portò al loro progressivo addomesticamento. È probabile che questi primi mandriani abbiano mantenuto uno stile di vita nomade o seminomade, mentre altri abitanti della Mezzaluna fertile (presumibilmente rappresentati dagli aplogruppi E1b1b, G e T) si sedentarizzarono e cominciarono a coltivare la terra o ad allevare animali più piccoli.
Lanalisi del DNA bovino rivela che tutti i bovini odierni (Bos taurus) discendono da una popolazione di soli ottanta uri ancestrali. Le prove più antiche della domesticazione dei bovini risalgono all8.500 a.C. circa e sono state trovate tra le culture del neolitico preceramico che popolavano i monti del Tauro. I due siti archeologici che per primi hanno mostrato tracce di bovini domestici sono i villaggi di Çayönü Tepesi nella Turchia sudorientale e Djade el-Mughara in Iraq settentrionale, separati da circa duecentocinquanta chilometri. Si presume che sia proprio questa larea da cui le linee R1b cominciarono a espandersi; in altre parole, questa sarebbe la terra dorigine di R1b.
I primi mandriani R1b si sarebbero successivamente separati in almeno tre gruppi. Un ramo (M335 o R1b2) si fermò in Anatolia ma, vista la sua scarsa incidenza odierna, non pare ebbe molto successo, forse a causa della forte competizione con le altre popolazioni dellAnatolia neolitica e alla mancanza di pascoli in questo territorio così impervio. Un secondo ramo sarebbe migrato verso sud, nel Levante, dove divenne il ramo V88. Alcuni di questi pastori avrebbero cercato nuove terre spingendosi ancora più a sud, in Egitto e poi in nord africa, dalla costa del Mediterraneo fino al Sahel. Il terzo ramo (P297) attraversò il Caucaso per poi spostarsi nelle vaste steppe pontico-caspiche, le quali rappresentavano il territorio ideale per le mandrie. Questi ultimi si divisero in due fazioni: R1b1a1 (M73), che migrò verso est in Asia centrale seguendo le coste del Mar Caspio, e R1b1a2 (M269) che in un primo tempo rimase nel Caucaso settentrionale e nelle steppe pontiche tra il Dnepr e il Volga. Non è chiaro se M73 raggiunse lAsia centrale attraversando il Caucaso e tagliando per il Kazakhstan, o se invece scelse la via meridionale che passa dallIran e dal Turkmenistan. In ogni caso, proprio come V88 e M335, M73 rappresenterebbe un ramo preindoeuropeo di R1b.
R1b-M269, la forma più comune in Europa, è strettamente associata alla diffusione delle lingue indoeuropee. Ciò del resto è attestato dalla sua presenza in tutte le regioni del mondo dove, già nellantichità, si parlavano queste lingue: dalla costa atlantica dellEuropa fino al subcontinente indiano; in sostanza in tutta lEuropa escluse la Finlandia, la Sardegna e la Bosnia-Erzegovina; lo si ritrova anche in Anatolia, in Armenia, nella Russia europea, in Siberia meridionale, in alcune sacche intorno allAsia centrale (Xinjiang, Turkmenistan, Tajikistan e Afghanistan), senza dimenticare lIran, il Pakistan, lIndia settentrionale e il Nepal. La storia di R1b e quella di R1a sono strettamente interconnesse.
Il ramo africano e levantino di R1b (V88)
Proprio come la sua controparte settentrionale (R1b-M269), anche R1b-V88 è associato alla domesticazione dei bovini, avvenuta per la prima volta in Mesopotamia settentrionale. È probabile che i rami si siano divisi subito dopo questo evento avvenuto allincirca 10.500 anni fa (8.500 a.C.). R1b-V88 migrò a sud verso il Levante e in Egitto, e questa migrazione può essere ricostruita archeologicamente seguendo le tracce dei bovini domestici, i quali apparvero nella Siria centrale intorno all8.000-7.500 a.C. (tardo periodo di Mureybat), e in Egitto e nel Levante meridionale intorno al 7.000-6.500 a.C. (per esempio a Nabta Playa e a Bir Kiseiba). Da qui i mandriani si diffusero nella quasi totalità dellAfrica settentrionale e orientale. Sembra che durante il Neolitico subpluviale (circa 7.250-3.250 a.C.) il deserto del Sahara fosse molto più umido di oggi e assomigliasse a una vasta savana ricoperta di erba, lambiente ideale per lallevamento dei bovini.
In Libia centrale, a Uan Muhuggiag, le prime testimonianze dellallevamento dei bovini risalgono al 5.500 a.C., mentre nelle grotte di Capelletti, in Algeria settentrionale, sono databili al 4.500 a.C. Tuttavia, le prove più evidenti delle relazioni tra gli Europei moderni e le genti che in passato abitarono il Sahara si trovano a Tassili n'Ajjer in Algeria meridionale, un sito famoso per i suoi petroglifi (incisioni rupestri) risalenti proprio al Neolitico. Alcune pitture sono databili al 3.000 a.C. e rappresentano donne bionde o fulve dalla pelle chiara nellatto di cavalcare delle mucche. Il campione R1b-V88 più antico mai ritrovato in Europa appartiene a un individuo di 6.200 anni fa, un allevatore-agricoltore della Catalogna, analizzato da Haak et al. (2015). Dal punto di vista autosomico questo individuo è un tipico agricoltore del Vicino Oriente: possiede, infatti, solo una piccolissima parte di miscela genetica Mesolitica dellEuropa occidentale.
Dopo aver raggiunto il Maghreb, i mandriani R1b-V88 potrebbero aver attraversato lo stretto di Gibilterra per spostarsi nella penisola Iberica, probabilmente accompagnati da contadini G2 e J1 e da pastori di capre T1a. Potrebbero essere stati proprio questi pastori-agricoltori neolitici ad aver dato inizio alla cultura della ceramica almagra nellAndalusia del VI millennio a.C.
Oggi, R1b-V88 si trova con basse frequenze (1-4%) tra i Libanesi, tra i drusi e tra gli Ebrei, così come in quasi tutti i paesi africani a nord dellequatore. Frequenze più elevate si possono riscontrare in Egitto (5%), tra i Berberi del confine libico egiziano (23%), tra i Sudanesi Copti (15%), tra le genti Hausa del Sudan (40%), tra i popoli Fulani del Sahel (54% in Niger e in Camerun), e tra le tribù ciadiche della Nigeria e del Camerun settentrionali (specialmente tra i Kirdi), dove si osservano frequenze che vanno dal 30 al 95%. Secondo Cruciani et al. (2010) R1b-V88 avrebbe attraversato il Sahara intorno ai 9.200 e i 5.600 anni fa. Con tutta probabilità sarebbe associabile alla diffusione delle lingue ciadiche, un ramo delle lingue afroasiatiche. V88 sarebbe poi migrato dallEgitto al Sudan, per poi espandersi lungo il Sahel fino al nord del Camerun e della Nigeria. In ogni caso, R1b-V88 non è presente solo tra i parlanti ciadico ma anche tra i parlanti senegambiano (Fulani-Hausa) e i parlanti lingue semitiche (Berberi e Arabi).
R1b-V88 si trova anche tra le popolazioni native del Ruanda, in Sudafrica, in Namibia, in Angola, in Congo, in Gabon, nella Guinea Equatoriale, nella Costa dAvorio, in Guinea Bissau. La sua ampia distribuzione in quasi tutte le parti del continente africano, la sua alta incidenza tra le tribù di mandriani e la sua età di coalescenza sono elementi a favore di una sua diffusione neolitica. In ogni caso, una migrazione più tardiva a partire dallEgitto sarebbe improbabile, poiché in questo caso insieme a V-88 dovremmo poter ritrovare quegli aplogruppi che apparvero in Egitto durante letà del bronzo, come J1, J2, R1a o R2b-L23.
Le linee materne associate alla diffusione di R1b-V88 in Africa sono gli aplogruppi mitocondriali J1b, U5, V e forse anche U3 e alcune sottocladi di H (=> vedi Analisi degli aplogruppi mitocondriali delle genti R1b originarie).
Il Caucaso settentrionale e le steppe pontico caspiche: il legame con gli Indoeuropei
I linguisti moderni hanno localizzato la terra natale dei protoindoeuropei nelle steppe pontico-caspiche, una precisa regione geografica e archeologica che si estende dallestuario del Danubio fino ai monti Urali a Est e fino al Caucaso settentrionale a sud. Le culture delle steppe pontico-caspiche del periodo Neolitico, dellEneolitico o della prima età del bronzo, cadono tutte sotto la denominazione di cultura Kurgan (4.200-2.200 a.C.) elaborata per la prima volta da Marija Gimbutas. Ciò è dovuto allusanza, mantenuta dalle diverse culture che hanno di volta in volta abitato la regione, di inumare i morti in tumuli chiamati appunto kurgan. È ormai noto che i siti di sepoltura di tipo kurgan apparvero a partire dal IV millennio a.C., e quasi certamente ebbero origine nel Caucaso meridionale. Dato che la variabilità genetica di R1b è massima intorno allarea orientale della penisola Anatolica, è molto difficile negare che questo aplogruppo si sia evoluto da queste parti prima di entrare nel mondo delle steppe.
I cavalli furono addomesticati per la prima volta intorno al 4.600 a.C. nelle steppe Caspiche, forse intorno al Don o al basso Volga, e presto divennero uno degli elementi caratteristici delle culture delle steppe. Comunque sia, è improbabile che in questo periodo R1b fosse già presente nelle steppe orientali, e dunque la domesticazione del cavallo andrebbe attribuita alle genti autoctone R1a, o alle tribù appartenenti al più antico ramo R1b-P297 che si stabilì in Europa orientale durante il tardo Paleolitico o durante il Mesolitico. Tutti i campioni provenienti dalla cultura mesolitica di Samara (Haak 2015) e dalla Lettonia (Jones 2017) appartengono al gruppo R1b-P297. Dal punto di vista autosomico questi individui mesolitici R1a e R1b erano Europei Mesolitici Orientali quasi puri, talvolta con una leggera miscela siberiana, ma mancavano del tutto della miscela caucasica ritrovata nei campioni provenienti dalle culture calcolitiche di Afanasevo, di Jamna e dalla cultura della ceramica cordata.
Non è ancora del tutto chiaro quando esattamente R1b-M269 passò dal Caucaso meridionale alle steppe pontico-caspiche. Potrebbe essere avvenuto in concomitanza con lemergere della cultura del Dnepr-Donec (circa 5.100-4.300 a.C.), che rappresenta la prima vera cultura delle steppe pienamente neolitica. Gli animali domestici (bovini, pecore e capre) erano allevati in tutte le steppe, e i rituali funebri erano molto elaborati. La lana delle pecore ricoprì un ruolo fondamentale nella società indoeuropea, influenzando labbigliamento soprattutto dei Celti e dei Germani (rami R1b degli Indoeuropei). In ogni caso, vi sono molti elementi che indicano una continuità tra la cultura del Dnepr-Donec e i cacciatori-raccoglitori mesolitici precedenti, e allo stesso tempo pare che questa cultura sia stata influenzata anche dalle culture balcaniche e dei Carpazi, da cui importava ceramiche e oggetti di rame. È probabile dunque che la cultura di Dnepr, più che uninvasione di R1b, marchi invece il periodo di transizione durante il quale le genti indigene R1a e/o I2a1b passarono a uneconomia agricola, forse anche grazie allinfluenza degli agricoltori del vicino Oriente già stanziatisi nella Vecchia Europa. I trenta campioni analizzati da Mathieson et al. (2017). provenienti dallUcraina neolitica (5.500-4.800 a.C.) appartengono agli aplogruppi I, I2a2, R1a, R1b1a (L754) e un R1b1a2 (L388). Nessuno appartiene a R1b-M269 o R1b-L23, cioè quelle cladi che dominarono durante il periodo Jamna. Anche le linee mitocondriali hanno unorigine esclusivamente Europea Mesolitica (U4a, U4b, U4d, U5a1, U5a2, U5b2 e un J2b1 e un U2e1). Nessuna di queste linee materne include aplogruppi tipicamente indoeuropei come H2a1, H6, H8, H15, I1a1, J1b1a, W3, W4 o W5, i quali apparvero più tardi nelle culture di Jamna, nella cultura della ceramica cordata e in quella di Unetice. Di conseguenza, dal punto di vista autosomico i genomi provenienti dallUcraina neolitica sono puri genomi Europei Mesolitici (per circa il 90% Cacciatori-Raccoglitori Orientali (EHG) e per il 10% Cacciatori-Raccoglitori Occidentali (WHG)) e mancano completamente della miscela Caucasica (CHG) che compare nella cultura di Jamna e nelle successive culture indoeuropee delletà del bronzo.
Le prime culture chiaramente protoindoeuropee furono le culture delle steppe ponto caspiche di Chvalynsk (5.200-4.500 a.C.) e di Srednij Stog (4.600-3.900 a.C.). È a questo punto che cominciarono ad apparire piccole sepolture di tipo kurgan, nelle quali i corpi venivano posti in posizione supina con le ginocchia piegate e orientate verso nordest, tratto che si trova anche nelle successive culture delle steppe. La varietà dei crani recuperati da queste tombe indica che ci fu una sovrapposizione di diverse popolazioni. Verso la fine del V millennio, iniziò a svilupparsi una élite che usava come simboli del proprio potere le enormi mandrie di bovini e di cavalli e il possesso di oggetti in rame. La principale sottoclade di R1b-M269, L23, sembra sia comparsa nel periodo di passaggio tra la cultura di Chvalynsk e quella di Srednij Stog intorno al 4.500 a.C. Il 99% degli Indoeuropei R1b discende da questa clade L23. Un altro ramo derivato da M269 è PF7562 che oggi si trova soprattutto nei Balcani, in Turchia e in Armenia e che rappresenterebbe una precoce migrazione proprio verso i Balcani intorno al periodo di Srednij Stog.
Appena prima del 3.700 a.C. ci fu una seconda migrazione attraverso il Caucaso, in concomitanza alla comparsa quasi improvvisa, nel Caucaso nordoccidentale, della prima società delletà del bronzo della storia: la cultura Majkop. Le origini di questa cultura sono ancora avvolte nelloscurità, ma gli archeologi hanno cercato di collegarla alle contemporanee culture calcolitiche dellAssiria e dellIran occidentale. Larcheologia ha rivelato anche che la lavorazione del bronzo e le sepolture kurgan si sarebbero diffuse dalla cultura Majkop verso le steppe pontiche, dove appena più tardi si sviluppò la cultura Jamna (dal 3.500 a.C.). I kurgan (o tumuli) sarebbero poi diventati la caratteristica dominante delle antiche società indoeuropee e furono ampiamente utilizzate dai Celti, dai Romani, dalle tribù germaniche e dagli Sciti.
Il periodo Jamna (3.500-2.500 a.C.) è un periodo fondamentale nella formazione della cultura e della società indoeuropea. Le genti mediorientali R1b-M269 per almeno un millennio, ma forse anche due o tre, hanno vissuto e si sono mischiate con i cacciatori raccoglitori e con i mandriani autoctoni R1a. La stretta vicinanza culturale e le interazioni tra R1a e R1b in tutte le steppe pontico-caspiche portarono alla nascita di una parlata comune, una nuova lingua franca, che i linguisti hanno chiamato protoindoeuropeo (PIE). Non ha alcun senso tentare di assegnare al PIE una diversa zona di origine. Esistono forti somiglianze linguistiche tra il PIE, le lingue urrite e caucasiche del Medioriente e le lingue uraliche della regione del Idel-Ural, e in questo caso le steppe pontiche sono lintermediario perfetto per collegare queste regioni.
È durante il periodo Jamna che i pastori-mandriani di pecore e bovini adottarono il carro per il trasporto del cibo e delle tende. Questa innovazione tecnologica permise loro di penetrare più in profondità nelle steppe, dando inizio a un nuovo stile di vita più mobile che a un certo punto sfociò nelle grandi migrazioni indoeuropee. Questo tipo di migrazioni di massa dove intere tribù si spostavano per mezzi di carri da trasporto erano praticate ancora al tempo di Giulio Cesare dai Galli, e dai Germani almeno fino alla tarda antichità.
Lorizzonte Jamna non rappresentò mai una singola cultura unificata. A sud, lungo le coste settentrionali del mar Nero fino al Caucaso nordorientale, vi era una regione di steppe aperte che si estendeva a est fino al mar Caspio, alla Siberia e alla Mongolia (le steppe eurasiatiche). La porzione occidentale tra il Don e il Dnestr (e più tardi fino al Danubio) era la regione più densamente abitata dalle genti R1b tra cui spiccava una minoranza (5-10%) di R1a. La porzione orientale nel bacino del Volga fino ai monti Urali, era al contrario abitata da genti R1a mischiata a una nutrita minoranza di R1b. I discendenti di questultimi si trovano ancora oggi tra i Baschiri, tra i Turkmeni, tra gli Uiguri e tra gli Hazara. La parte settentrionale dellarea Jamna era costituita da steppe-foreste occupate da genti R1a, anchesse unite a una piccola porzione di R1b (almeno a giudicare dai campioni provenienti dalla cultura della ceramica cordata e dalla Russia e dalla Bielorussia moderne, dove la frequenza di R1b è dalle sette alle nove volte più bassa di R1a). Sarebbe stato il ramo occidentale a migrare, in ondate successive, prima nei Balcani e in Grecia, poi in Europa centrale e occidentale, per poi tornare indietro nella propria patria ancestrale in Anatolia (Ittiti, Frigi, Armeni ecc.). Il ramo orientale sarebbe invece migrato in Asia centrale, nello Xinjiang, in Siberia e in Asia meridionale (Iran, Pakistan, India). Il ramo settentrionale si sarebbe evoluto nella cultura della ceramica cordata che si diffuse intorno al Baltico, la Polonia, la Germania e la Scandinavia.
La cultura Majkop rappresenta il legame tra R1b e le steppe?
La cultura di Majkop (3.700-2.500 a.C.) del Caucaso nordorientale fu, dal punto di vista culturale, una sorta di estensione dellorizzonte Jamna. Nonostante non sia normalmente considerato parte delle culture delle steppe pontico-caspiche, in parte proprio per la sua localizzazione geografica, il Caucaso settentrionale ebbe legami fortissimi con le steppe, come attestato da numerose ceramiche, dalloro, dalle armi in rame e in bronzo e dai gioielli delle contemporanee culture di Mikhaylovka, di Srednij Stog e di Kemi Oba. In realtà la costa settentrionale del mar Nero e il nord del Caucaso erano in contatto già prima del periodo Majkop. Lantenata del periodo Majkop, la cultura di Svobodnoe (4.400-3.700 a.C.), ebbe infatti diversi scambi con le culture Suvorovo-Novodanilovka e di Srednij Stog. Ancora più antichi, gli insediamenti di Nalchik (5.000-4.500 a.C.) nel Caucaso settentrionale presentano una cultura simile a quella di Chvalynsk delle steppe caspiche e della regione del Volga. Potrebbe essere proprio questo il periodo durante il quale gli R1b cominciarono a interagire e a mescolarsi con le popolazioni R1a delle steppe.
Sia le genti Jamna che le genti Majkop utilizzavano i kurgan come siti di sepoltura, e inumavano i loro defunti in posizione supina con le ginocchia alzate e orientate lungo lasse nordest/sudovest. I pavimenti delle tombe erano cosparsi di ocra rossa, e alcuni animali domestici venivano sacrificati e seppelliti accanto agli esseri umani. Altri tratti che accomunano le due culture sono: lutilizzo del cavallo e del carro; uneconomia dipendente dalle mandrie di bovini e dalle greggi di pecore, utilizzate soprattutto per la lana; luso di asce da battaglia in rame o in bronzo e di daghe dotate di codolo.
I primi carri e utensili in bronzo della storia sono stati ritrovati nel Caucaso settentrionale, e sembra si siano diffusi nelle steppe proprio a partire da questa regione.
Quella di Majkop fu una delle culture più avanzate delletà del bronzo, in verità una delle prime a sviluppare la metallurgia e di conseguenza ad avere armi in metallo. La più antica spada in bronzo è stata ritrovata in una tomba del tardo periodo Majkop, nel kurgan 31 di Klady. Ricorda nello stile le lunghe spade celtiche, nonostante sia meno elaborata. I resti e le raffigurazioni dei cavalli che compaiono già nelle prime tombe Majkop suggeriscono che allorigine di questa cultura potrebbero esserci genti delle steppe o popolazioni a esse legate. In ogni caso, la presenza anche di elementi culturali radicalmente differenti da quelli delle culture delle steppe, almeno in alcuni siti, starebbe a significare che quella di Majkop fu una popolazione ibrida. Purtroppo, senza campioni di DNA antico da analizzare, è impossibile dire se la cultura di Majkop si sia formata solo grazie alla sovrapposizione di gruppi anatolici R1b e di gruppi caucasici G2a, o se si siano aggiunte anche delle tribù R1a. Queste due o tre etnie potrebbero aver coabitato fianco a fianco in diversi insediamenti. Lunica linea di DNA Y tipicamente caucasica la cui distribuzione ricalca il pattern delle migrazioni indoeuropee è G2a-L13, che si trova in tutta lEuropa e in Asia centrale e meridionale. Nei Balcani, nel bacino del Danubio e in Europa centrale la sua frequenza è proporzionale alla frequenza di R1b.
Si ritiene che le genti Majkop siano state le prime a introdurre nelle steppe, a partire dalla Mesopotamia, lutilizzo di veicoli primitivi dotati di ruote (carri). Ciò avrebbe rivoluzionato lo stile di vita di chi abitava questi luoghi, e intorno al 2.000 a.C avrebbe portato allinvenzione del carro da guerra (trainato da cavalli). La cavalleria e i carri giocarono un ruolo fondamentale nelle successive migrazioni indoeuropee, permettendo spostamenti così rapidi da determinare la sconfitta di chiunque si trovassero di fronte. Se si combinano queste tecnologie con le più avanzate armi in bronzo e la dimestichezza di questi popoli con il mare e la navigazione, il ramo occidentale (R1b) degli Indoeuropei proveniente dalle coste del mar Nero diviene un eccellente candidato per identificare i misteriosi Popoli del Mare che devastarono le coste orientali del Mediterraneo durante il II millennio a.C.
Gli Ittiti, che parlavano una lingua indoeuropea, comparvero in Anatolia centrale appena qualche secolo dopo la scomparsa delle culture di Majkop e di Jamna. Se si considera che la maggior parte delle forme indoeuropee di R1b oggi presenti in Anatolia appartengono alla sottoclade R1b-Z2103, è quasi certo che gli Ittiti arrivarono in Anatolia attraverso i Balcani, proprio dopo che le genti Jamna/Majkop invasero lEuropa sudorientale. A queste due culture seguì la cultura di Srubna (1.600-1.200 a.C.), la quale rappresenterebbe lavanzata dei popoli R1a-Z282 dalle steppe settentrionali verso le coste del mar Nero, la quale mirò a riempire il vuoto lasciato dalle tribù R1b ormai emigrate in Europa sudorientale e in Anatolia.
 - Eupedia")
Mappa delle migrazioni dellaplogruppo R1b nel tardo Neolitico e nella prima età del bronzo (5.500-3.000 a.C.)
Il ramo siberiano e centroasiatico
Quando, nel tardo neolitico, R1b attraversò il Caucaso, questa linea si divise in due gruppi principali. Il ramo occidentale (L51) si sarebbe stabilito nelle regioni a est e a nord del mar Nero. Il ramo orientale (Z2103) invece sarebbe migrato nella regione del Volga-Don dove, verso il 4.600 a.C.,. erano stati addomesticati i cavalli. Qui è molto probabile che R1b si sia mischiato alle genti indigene R1a per poi dare origine alla cultura Repin (3.700-3.330 a.C.), poco prima che comparisse la cultura di Jamna nelle steppe pontico-caspiche occidentali. R1b sarebbe poi migrato, grazie anche ai cavalli, attraverso le grandi steppe eurasiatiche, fino a raggiungere i monti Altaj nella parte orientale dellAsia centrale dove diede origine alla cultura di Afanasevo (circa 3.600-2.400 a.C.). Le genti Afanasevo potrebbero essere i precursori del ramo tocario delle lingue indoeuropee. Nel 2014, Clément Hollard dellUniversità di Strasburgo ha analizzato tre campioni di DNA Y provenienti dalla cultura di Afanasevo, e tutti e tre sono risultati appartenerne allaplogruppo R1b, inclusi due R1b-M269.
Le genti R1b che rimasero nella regione Idel-Ural furono probabilmente i precursori della cultura di Poltavka (2.700-2.100 a.C.), i quali in seguito si integrarono nella cultura di Sintashta-Petrovka (2.100-1.750 a.C.) a maggioranza R1a e legata alla conquista indoariana dellAsia centrale e meridionale (=> vedi R1a per altri dettagli).
Oggi, in Russia, R1b è più diffuso tra le minoranze etniche della regione dellIdel-Ural (Udmurti, Komi, Mordvini, Tartari) che tra i Russi di origine slava. R1b è presente anche tra diverse popolazioni dellAsia centrale, e in questo caso le percentuali più alte si osservano tra gli Uiguri (20%) dello Xinjiang nel nordovest della Cina, tra gli Yagnobi del Tajikistan (32%) e tra i Baschiri 47% o 62.5% nel distretto di Abzelilovsky) di Bashkortostan in Russia (ai confini con il Kazakistan).
R1b-M73 si trova principalmente in Asia settentrionale (Altaj, Mongolia), in Asia centrale e nel Caucaso settentrionale. Si ritiene si sia diffuso dal Medioriente durante il Neolitico da dove si sarebbe spostato verso lAsia centrale e settentrionale, e può dunque essere considerato preindoeuropeo.
Il ramo europeo e mediorientale
Le armi in bronzo e la formidabile mobilità provvista dai cavalli avrebbero fornito agli Indoeuropei un considerevole vantaggio sulle popolazioni autoctone europee per lo più costituite dagli aplogruppi C1a2, F e I (discendenti dei Cro-Magnon) e dai pastori/agricoltori neolitici G2a, H2, E1b1b e T1a. Questo permise a R1a e a R1b di rimpiazzare abbastanza rapidamente la maggior parte delle linee maschili native (=>vedi Come furono rimpiazzate le linee più antiche dellEuropa occidentale dallarrivo di R1b?), mentre le linee femminili pare siano state meno coinvolte da questo processo.
Se si analizza linvasione indo-iraniana dellAsia meridionale si scopre che, oggi, il 40% delle linee maschili dellIndia settentrionale è composto da R1a, ma meno del 10% delle linee femminili sembra avere origini indoeuropee. In Europa, limpatto degli Indoeuropei fu ancora più forte perché, rispetto alla civiltà della Valle dellIndo. Le società europee di 4.000 anni fa erano molto meno sviluppate sia dal punto di vista delle tecniche agricole che della tecnologia militare (non avevano armi in bronzo) e della densità di popolazione. Questo è vero soprattutto per le popolazioni dellEuropa occidentale, dove lagricoltura arrivò molto più tardi rispetto ai Balcani o allEuropa centrale. In Grecia, nei Balcani e nei Carpazi vi erano le più avanzate società europee del tempo ed esse furono meno soggette al rimpiazzamento genetico. Le linee neolitiche sopravvissero meglio anche in quelle regioni più difficili da raggiungere o meno ospitali per gli allevatori di cavalli, come le Alpi, le Alpi dinariche, gli Appennini e la Sardegna.
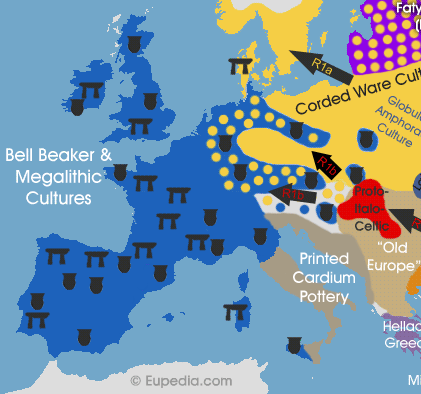
La conquista della Vecchia Europa e dellEuropa centrale (4.200-2.500 a.C.)
Le prime incursioni dei popoli delle steppe nei Balcani avvennero tra il 4.200 e il 3.900 a.C., quando i mandriani di bovini, equipaggiati con i loro carri trainati da cavalli, attraversarono il Dnestr e il Danubio per poi distruggere le città delle culture di Gumelnita, di Varna e di Karanovo VI in Romania orientale e in Bulgaria. Probabilmente fu un cambiamento climatico, che proprio in quel periodo rese gli inverni più rigidi, a spingere i mandriani delle steppe in cerca di pascoli più verdi, mentre diverse carestie portavano alla fame e allinstabilità le comunità danubiane e balcaniche. Le successive culture di cultura di Cernavodă (calcolitico 4.000-3.200 a.C.), di Coțofeni/Usatovo (età del rame/età del bronzo 3.500-2.500 a.C.) e di Ezero (età del bronzo 3.300-2.700 a.C.), tutte originarie dellodierna Romania, sembra siano il risultato della sovrapposizione di genti che già abitavano gli antichi insediamenti di tipo tell e dei nuovi immigrati arrivati dalle steppe. Questi immigrati portarono con tutta probabilità un insieme di linee R1a e R1b, forse con una percentuale di R1a più alta rispetto alle più tarde invasioni dellera Jamna.
Gli invasori provenienti dalle steppe avrebbero forzato gli agricoltori danubiani a migrare verso gli insediamenti della cultura di Cucuteni-Trypillian nei Carpazi orientali, generando ivi un boom demografico che portò questultima a espandersi verso nordest fino alla valle del Dnepr, spingendo fino allUcraina centrale gli aplogruppi-Y G2a, I2a1 (probabilmente la linea dominante della cultura di Cucuteni-Trypillian), E1b1b, J2a e T1a. Questa precoce avanzata indoeuropea verso ovest fu piuttosto limitata, anche perché le armi in bronzo non erano ancora completamente diffuse e mancavano gli eserciti organizzati. Essa fu possibile solo grazie ai cambiamenti climatici che avevano ridotto le difese degli insediamenti della Vecchia Europa. Le culture dei Carpazi, del Danubio e dei Balcani erano troppo popolose e tecnologicamente avanzate per permettere una migrazione di massa.
Dallaltra parte, le genti R1a delle steppe-foreste penetrarono con successo nel cuore dEuropa incontrando ben poca resistenza, facilitate dallassenza di società agricole sviluppate tra la Polonia e il Baltico. La cultura della ceramica cordata (3.200-1.800 a.C.) rappresentò la naturale espansione, verso nord e verso ovest, della cultura di Jamna, e raggiunse da una parte la Germania e dallaltra la Svezia e la Norvegia. Lanalisi dei campioni di DNA appartenenti alla cultura della ceramica cordata ha confermato la presenza di R1a e R1b in Polonia già intorno al 2.700 a.C., e di R1a in Germania centrale intono al 2.600 a.C. Queste tribù si espansero a partire dal margine settentrionale della cultura di Jamna, dove le linee R1a dominavano su quelle R1b.
Come si diceva, in Europa, lespansione delle genti R1b fu molto più lenta, ma alla fine anche qui si rivelò inesorabile. Verso il 2.800 a.C., quando la ceramica cordata aveva già raggiunto la Scandinavia, le culture R1b delletà del bronzo avevano appena mosso i primi passi nelle steppe della Pannonia. Esse fondarono importanti insediamenti nella grande pianura ungherese, che offriva lhabitat più simile alle ancestrali steppe pontiche da cui erano partiti millenni addietro. Intorno al 2.500 a.C., il ramo occidentale degli Indoeuropei R1b era pronto per la successiva migrazione di massa verso la Germania odierna e verso lEuropa orientale. A quel punto, gli immigranti R1b si erano già ampiamente mischiati con le popolazioni mesolitiche e neolitiche autoctone del bacino del Danubio, dove ormai avevano convissuto per 1.700 anni.
Lélite indoeuropea, senza dubbio patriarcale, dal lato maschile rimase composta quasi esclusivamente da R1b ma assorbì una grande porzione di linee materne non indoeuropee. Così ibridizzate, le nuove genti protoindoeuropee R1b persero quasi tutti i loro tratti protoeuropoidi o mongolidi, eredità delle loro origine caspiche (che erano ancora visibili in numerosi individui del periodo Jamna). I loro capelli, la pigmentazione degli occhi e della pelle originariamente più chiari, una volta mescolatisi con i più scuri abitanti della Vecchia Europa divennero più simili a quelli dei moderni Europei meridionali. Le genti R1a della cultura della ceramica cordata, invece, spostandosi in Europa settentrionale vennero in contatto con società molto meno avanzate e meno popolose, per lo più discendenti dalle più chiare popolazioni mesolitiche. Mantennero dunque con più facilita la loro pigmentazione originaria. In ogni caso, anche tra gli R1a della Scandinavia i tratti del viso si evolvettero considerevolmente, poiché in quellarea gli abitanti I1 erano per lo più dolicocefali dai visi allungati, lopposto dei brachicefali dai visi larghi provenienti dalle steppe.
La conquista dellEuropa occidentale (2.500-1.200 a.C.)
La conquista dellEuropa da parte degli R1b avvenne in due fasi. Per circa due millenni, a partire più o meno dal 4.200 a.C., le genti delle steppe limitarono le loro conquiste alle ricche civiltà calcolitiche dei Carpazi e dei Balcani. Di queste società facevano parte i più grandi insediamenti del tempo, in particolare gli insediamenti di tipo tell della cultura di Cucuteni-Trypillian. Durante questa prima fase non cera niente che potesse spingere i conquistatori R1b a muoversi ancora più a occidente, poiché la maggior parte delle terre a nord e a ovest delle Alpi erano ancora scarsamente abitate e ricoperte di foreste. Il neolitico non raggiunse le isole Britanniche e la Scandinavia prima del 4.000 a.C. Anche nella Francia settentrionale, e lungo quasi tutto larco alpino, lagricoltura esisteva da poco meno di un millennio ed era ancora primitiva, se comparata a quella dellEuropa sudorientale e del Medioriente.
Per secoli, dopo la conquista dei Balcani da parte degli Indoeuropei, il resto dellEuropa nordoccidentale rimase ancora abitata da società tribali di cacciatori-raccoglitori che praticavano solo sporadicamente lagricoltura. Perché mai i nostri conquistadores R1b avrebbero dovuto lasciare le comodità delle ricche e popolose civiltà danubiane per le dure condizioni di vita che li aspettavano a occidente? Inoltre, le genti delletà del bronzo cercavano soprattutto stagno, rame e oro di cui i Balcani erano ricchi, ma che ancora non erano stati scoperti in Europa occidentale.
Pare che R1b-L51 sia arrivato in Europa centrale (Ungheria, Austria, Boemia) intorno al 2.500 a.C., più o meno due millenni dopo linizio della rivoluzione neolitica in queste regioni. Qui gli insediamenti agricoli avevano appena cominciato a svilupparsi, e lestrazione di rame e oro era agli albori. La prospettiva di una conquista di questi territori non era particolarmente allettante.
Le testimonianze archeologiche e le prove genetiche (come la distribuzione delle sottocladi di R1b) indicano che, tra il 2.800 e il 2.300 a.C., la Germania centrale e orientale furono investite da una serie di ondate migratorie consecutive. La cultura di Unetice fu probabilmente la prima cultura dove le linee R1b-L11 giocarono un ruolo fondamentale. È interessante notare come il periodo di Unetice cominci in corrispondenza della fine delle culture di Majkop (2.500 a.C.) e di Kemi Oba (2.200 a.C.) sulle coste settentrionali del mar Nero, quando queste furono rimpiazzate da culture originarie delle steppe settentrionali. Si può perciò supporre che le popolazioni a maggioranza R1b provenienti dalla metà settentrionale del mar Nero siano migrate verso occidente a causa della pressione esercitata da altre genti indoeuropee (R1a) provenienti da nord, per esempio quelle del nascente ramo protoindoiraniano, collegato alle contemporanee culture di Poltavka e Abashevo.
È poco probabile che la cultura del vaso campaniforme (2.800-1.900 a.C.) dellEuropa occidentale rappresenti una cultura già indoeuropea, poiché le sue caratteristiche sono in perfetta continuità con le precedenti culture megalitiche locali. Il fenomeno dei vasi a campana cominciò in Portogallo tra la fine del neolitico e il calcolitico, e da qui si propagò in direzione nordest verso la Germania. Durante lo stesso periodo, le culture delle steppe delletà del bronzo si diffusero nella direzione opposta dalla Germania verso la penisola Iberica, verso la Francia e verso la Gran Bretagna, portando progressivamente le linee R1b nei territori del vaso campaniforme. È più probabile che questo tipo di ceramica e i cavalli ritrovati in tutta lEuropa occidentale risalenti a questo periodo siano il risultato di scambi commerciali con le vicine culture indoeuropee, incluse quelle della prima ondata R1b in Europa centrale. È altresì possibile che le genti del vaso campaniforme fossero in realtà mercanti o esploratori R1b che, dopo aver viaggiato attraverso lEuropa occidentale, riportarono indietro racconti sulle ricchezze mal difese da quelle genti delletà della pietra; ricchezze che non aspettavano altro che essere conquistate. Questo avrebbe favorito uninvasione su larga scala della Germania da parte dei popoli indoeuropei (R1b) a partire da circa il 2.500 a.C.. Essa avrebbe raggiunto lAtlantico (almeno a nord dei Pirenei) verso il 2.200 a.C.
Le analisi di antichi campioni di DNA condotte da Lee et al. (2012), Haak et al. (2015) e Allentoft et al. (2015) hanno tutte confermato la presenza in Germania di R1b-L51 e delle sue sottocladi più profonde, come P312 e U152, a partire almeno dal periodo della cultura del vaso campaniforme. Tuttavia non hanno identificato nessun R1b proveniente dalle culture più antiche. I campioni tedeschi R1b della cultura del vaso campaniforme presentano solo circa il 50% di miscela autosomica Jamna e spesso possiedono DNA mitocondriali (mtDNA) neolitici non provenienti dalle steppe. Questo confermerebbe che gli invasori R1b, avanzando verso ovest, presero in mogli donne locali. Un altro studio di Olalde et al. (2017) ha confermato che gli esponenti iberici della cultura del vaso campaniforme erano geneticamente distinti dagli individui tedeschi. Nessuno degli individui spagnoli o portoghesi associati a questa cultura presenta miscele tipiche delle steppe, e nessuno appartiene allaplogruppo indoeuropeo R1b-L23 né alle sue sottocladi. Sembrano invece tutti appartenere alle tipiche linee megalitiche G2a, I2a1, I2a2 e a R1b-V88 di origine neolitica. Questo lavoro ha confermato anche lalta incidenza in Europa centrale di linee R1b-L51 durante il periodo del vaso campaniforme. In Gran Bretagna, gli individui megalitici appartenevano esclusivamente allaplogruppo I2 (per lo più I2a2 e I2a1-L161), ma furono interamente rimpiazzati da R1b-L51 (soprattutto dalla sua sottoclade L21) durante la prima età del bronzo. Ciò significa che la cultura del vaso a campana non sarebbe associabile a un particolare gruppo etnico. La ceramica campaniforme ebbe origine nella penisola Iberica megalitica, ma si espanse in seguito in Francia e in Europa centrale e fu adottata anche dalle tribù di invasori R1b-L51 provenienti dalle steppe. Questi la portarono con loro nelle isole Britanniche spazzando via la maggior parte delle popolazioni megalitiche indigene. Quando si studia la cultura del vaso campaniforme, dunque, non si può parlare di una precisa popolazione, quanto piuttosto di varie popolazioni che durante questo periodo commerciarono e usarono ceramiche di questo tipo.
I campioni tedeschi di DNA della cultura di Unetice 2.300-1.600 a.C.), emersa nemmeno due secoli dopo che i primi individui R1b-L51 ebbero fatto la loro comparsa nella Germania della fine della cultura del vaso campaniforme, possiedono una percentuale leggermente più alta di ascendenza Jamna (60-65%) e di linee mitocondriali collegate a questa cultura. Ciò starebbe a indicare che la migrazione dalle steppe coinvolse sia gli uomini che le donne. Una migrazione spiegherebbe come mai i reperti archeologici della cultura di Unetice siano chiaramente connessi alla cultura di Jamna (cioè agli Indoeuropei), e spiegherebbe limprovvisa introduzione di nuove tecnologia e nuovi stili di vita, mentre la cultura del vaso campaniforme appare in continuità con le precedenti culture neolitiche o calcolitiche. Gli uomini R1b potrebbero aver conquistato queste terre limitandosi semplicemente a rovesciare i signori locali, senza per questo cancellare completamente la cultura autoctona, soprattutto a causa della loro inferiorità numerica. Facendo unanalogia con le migrazioni germaniche della tarda antichità, linvasione R1b del periodo del vaso a campana fu simile a quella di Goti, Burgundi e Vandali, i quali migrarono in piccoli gruppi creando nuovi regni allinterno dellImpero Romano, ma adottarono la lingua e la cultura romana. Al contrario, le culture della ceramica cordata e di Unetice furono il risultato di ampi flussi migratori delle genti delle steppe, le quali imposero la loro lingua indoeuropea e la loro cultura ai popoli conquistati, proprio come fecero gli Anglosassoni o i Bavari nel V secolo d.C.

| Argomenti correlati |
|---|
|
|
Le culture che succedettero alla cultura di Unetice in Europa centrale, in ordine cronologico, sono la cultura dei tumuli (1.600-1.200 a.C.), la cultura dei campi durne (1.300-1.200) e la cultura di Hallstatt (1.200-750 a.C.). Esse rimasero fondamentalmente indoeuropee. La cultura di Hallstatt, localizzata intorno alle Alpi, è considerata la prima cultura celtica classica dellEuropa. Essa si espanse rapidamente in Francia, in Gran Bretagna, nella penisola Iberica, in Italia settentrionale e nella valle del Danubio, probabilmente diffondendo per la prima volta le lingue celtiche, ma non la metallurgia del bronzo né le linee R1b che si erano già diffuse in quasi tutta lEuropa occidentale durante il periodo del vaso campaniforme.
(Vedi anche Come lestrazione del metallo e lallevamento di bestiame spiegano la dominanza di R1b sulla costa atlantica).
| Gli Indoeuropei hanno davvero invaso lEuropa occidentale? |
|---|
|
PChi, tra gli studiosi, propone un modello che presuppone una continuità tra le culture delletà del bronzo e quelle paleolitiche o neolitiche tende a sostenere che la metallurgia del bronzo e i cavalli siano stati importati dagli Europei occidentali grazie ai contatti con i loro vicini orientali, e che quindi non ci sia mai stata una vera e propria invasione indoeuropea. In questo caso però è difficile comprendere come le lingue italiche, celtiche e germaniche siano state adottate dagli Europei occidentali e settentrionali, se non si accetta nemmeno la possibilità di uninvasione su piccola scala. È stato suggerito che le lingue indoeuropee (IE) si siano diffuse attraverso i contatti tra diversi popoli, proprio come accade per certe tecnologie, forse perché rappresentavano la lingua di una piccola élite, e quindi la loro adozione avrebbe conferito un certo prestigio a chi le parlava. Eppure le persone non cambiano la propria lingua così facilmente, solo perché la nuova suona meglio o dona prestigio. Anche oggi, con laiuto di libri di testo, dizionari, corsi obbligatori di lingua, scuole private per adulti e programmi televisivi multilingua, la maggioranza delle persone fatica ad apprendere una lingua straniera appartenente a una famiglia completamente diversa dalla propria. La distanza linguistica tra le lingue preindoeuropee e quelle IE è comparabile più o meno a quella esistente tra linglese moderno e il cinese. Linglese, il greco, il russo e lhindi sono tutte lingue IE imparentate tra loro, e dunque sono più facili da imparare per un parlante IE che non le lingue non-IE come il cinese, larabo o lungherese. Dal punto di vista linguistico, solo una migrazione di parlanti IE su vasta scala potrebbe spiegare la capillare diffusione di queste lingue in Europa occidentale, migrazione che ha lasciato il basco come unico erede delle lingue neolitiche.
Unargomentazione, derivata da prove archeologiche, in favore della teoria della sostituzione etnica dei parlanti lingue neolitiche da parte di una cultura indoeuropea delletà del bronzo ci viene dallanalisi degli stili della ceramica. Limprovvisa comparsa della metallurgia del bronzo in Europa occidentale coincide con la diffusione di una ceramica più semplice e meno decorata, in tutto e per tutto simile a quella delle steppe pontiche. Per circa tremila anni, la ceramica si era costantemente evoluta verso una maggiore complessità e un maggior dettaglio. Non è credibile pensare che le popolazioni dellepoca abbiano deliberatamente deciso di ritornare verso uno stile più primitivo. È possibile che a un certo punto una qualche cultura isolata abbia deciso di sperimentare con una ceramica più semplice, ma quali sono le probabilità che culture distanti come quelle iberiche, galliche, italiche e britanniche abbiano tutte deciso di adottare questo nuovo stile, e per giunta nello stesso periodo? La spiegazione migliore è che questo nuovo stile sia stato imposto da invasori stranieri. In questo caso non è mera speculazione; ci sono numerose prove che la ceramica semplificata sia una caratteristica delle steppe associata allemergere dei protoindoeuropei.
Oltre alla ceramica, larcheologia ci fornisce numerose prove anche di un cambiamento nei modi di produzione di cibo. proprio durante la prima età del bronzo dellEuropa centrale e occidentale. Il peso dellagricoltura fu improvvisamente ridotto in favore dellallevamento. È sempre in questo periodo che si diffusero definitivamente i cavalli e si cominciò a consumare regolarmente il latte di mucca. Presi tutti insieme, questi cambiamenti ricalcano quasi perfettamente lo stile di vita delle steppe. Anche in seguito allintroduzione dellagricoltura, avvenuta intorno al 5.200 a.C., la cultura del Bug-Dnestr e le successive culture delle steppe furono caratterizzate da uneconomia dominata dalla pastorizia, dove lagricoltura rimaneva in secondo piano. Questo stile di vita si è diffuso in Europa proprio contemporaneamente alla lavorazione del bronzo.
Durante letà del bronzo, in Europa, anche le credenze religiose e le arti andarono incontro a un radicale cambiamento. Le società neolitiche europee e del Vicino Oriente veneravano da sempre delle figure femminili intese come simboli di un culto della fertilità. Con lespandersi della tecnologia del bronzo, dalla valle del Danubio verso lEuropa occidentale, i simboli della fertilità sparirono progressivamente e furono rimpiazzati da sculture di animali domestici.
Un altro indizio dellarrivo in massa degli Indoeuropei delle steppe si trova nelle pratiche di sepoltura. Gli Europei neolitici cremavano i propri morti (per esempio la cultura di Cucuteni-Trypillian) oppure li seppellivano in tombe collettive (per esempio le culture megalitiche). Nelle steppe invece, ogni persona era inumata individualmente, e le tombe di chi apparteneva agli strati sociali più alti erano poste in camere funerarie ricoperte da tumuli circolari. I corpi erano accompagnati da armi (mazze, asce, daghe), da ossa di cavalli e da un carro (più tardi un carro da guerra) smontato. Questi caratteristici tumuli funerari delle steppe pontiche sono conosciuti come kurgan. Gli uomini erano inumati in tombe più sontuose rispetto alle donne, anche se si trattava di bambini, e osservando le sepolture è facile identificare le differenze e le gerarchie sociali. Gli Indoeuropei vivevano in una società fortemente patriarcale e gerarchica, tutto lopposto delle culture matriarcali e più egalitarie della vecchia Europa. La proliferazione di questi kurgan nellEuropa centrale delletà del bronzo, i quali tenevano conto del grado sociale e del sesso del defunto, è un chiaro segnale del passaggio del potere a una élite indoeuropea. Queste pratiche si diffusero anche in Asia centrale e in Siberia meridionale, due regioni dove troviamo anche oggi le linee R1a e R1b. Tra tutti i riti che caratterizzano una cultura, quelli funerari sono tra i più significativi dal punto di vista emotivo, e più carichi di risvolti personali. È molto difficile che le persone cambino le loro tradizioni solo perché lo fanno i vicini. Non per niente già durante il neolitico e il calcolitico europeo coesistevano, fianco a fianco, diverse pratiche funerarie. Dunque leredità europea di unusanza tipica delle culture delle steppe pontico caspiche, e in questo caso di una tradizione che, abbiamo visto, non muta facilmente solo attraverso il contatto con i propri vicini, rafforza la tesi di una migrazione indoeuropea. Ladozione di alcuni tratti di una cultura straniera in genere avviene quando, grazie alla sua superiorità culturale, una civiltà schiaccia le culture adiacenti. Questo processo si chiama acculturazione. Non cè niente che indichi una superiorità culturale delle culture delle steppe tanto marcata da giustificare labbandono dei simboli fondamentali della propria cultura e della propria lingua da parte di popolazioni distanti dalle steppe più di duemila chilometri, come per esempio le culture atlantiche. In verità la Vecchia Europa era molto più avanzata nella produzione di ceramiche e gioielli. La superiorità indoeuropea non era culturale ma militare, ed era dovuta ai cavalli, alle armi in bronzo e a un codice etico che premiava gli atti di eroismo individuali in guerra (questi valori sono noti anche grazie agli antichi testi IE come i Rig Veda, gli Avesta o la letteratura micenea e ittita).
Dopo la linguistica e larcheologia, la terza disciplina che fornisce prove a favore di una migrazione è la genetica stessa. Allinizio si era ipotizzato che R1b fosse un aplogruppo nativo dellEuropa occidentale, perché è qui che oggi prevale. È stato invece dimostrato che, rispetto allEuropa, la maggiore variabilità nelle sequenze microsatellite degli aplotipi R1b si trova in Anatolia e nel Caucaso. Inoltre le sottocladi europee sono più recenti rispetto a quelle mediorientali o centroasiatiche. La clade principale europea, R-P312/S116, risale solo al 3.500-3.000 a.C. Ciò non significa che il più antico progenitore comune di questa linea non possa essere arrivato in Europa prima di questo periodo, ma significa che la prima persona portatrice della mutazione R-P312/S116 visse probabilmente nel basso Danubio o intorno al Mar Nero almeno 5.000 anni fa. In ogni caso, questa finestra temporale è troppo recente per poter dare sostegno alla tesi di unorigine di R1b paleolitica o neolitica. La scoperta di quelle che si pensavano essere linee europee in Asia centrale, in Pakistan e in India ha dato il colpo di grazia alla teoria dellorigine paleolitica europea di R1b e ha confermato il legame con gli Indoeuropei.
Tutti gli elementi concorrono in favore di una migrazione su larga scala di parlanti indoeuropeo (forse a grazie al cavallo) verso lEuropa occidentale tra il 2.500 e il 2.100 a.C. Questa contribuì al rimpiazzo dello stile di vita neolitico, o calcolitico, da parte di una cultura inerentemente delletà del bronzo, con una ceramica più semplice, meno fondata sullagricoltura e più sullallevamento, con nuove tradizioni (tombe singole) e nuovi valori (società patrilineare, eroi guerrieri) e che non si evolse a partire dai suoi predecessori locali.
|
Il ramo celtico atlantico (L21)
Le genti R1b proto-italo-celto-germaniche nel 2.500 a.C. erano già giunte in quella che oggi è la Germania. Verso il 2.300 a.C. il loro numero crebbe e nacque la cultura di Unetice. Seguendo il propagarsi della metallurgia del bronzo in Europa occidentale, pare che questi primi Indoeuropei abbiano raggiunto la Francia e i Paesi Bassi intorno al 2.200 a.C., la Gran Bretagna verso il 2.100 a.C., lIrlanda intorno al 2.000 a.C. e la penisola Iberica nel 1.800 a.C. È probabile che sia stata questa prima ondata di R1b a portare in queste regioni numerose linee R1b-L21 (forse a causa di un effetto del fondatore)), dato che queste sottocladi si trovano ovunque in Europa centrale, settentrionale e occidentale. Cassidy et al. (2015) hanno confermato la presenza di R1b-L21 (sottocladi DF13 e DF21) in Irlanda intorno al 2.000 a.C. Dal punto di vista autosomico, questi genomi assomigliano molto a quelli della cultura di Unetice e sono invece molto diversi da quelli del neolitico irlandese. Questo conferma che lintroduzione del bronzo in Irlanda avvenne ad opera di una migrazione diretta di R1b-L21 dallEuropa centrale.
Una separazione precoce di L21 dal principale ramo protoceltico, avvenuta intorno alla Germania, spiegherebbe perché le lingue celtiche Q (Goidelic and Hispano-Celtic) (goideliche e ispanoceltiche) siano così tanto diverse dal ramo delle lingue celtiche P (La Tène, gallico, brittonico), che pare si siano diffuse grazie alle successive culture di Hallstatt e dei campi durne.
Più tardi, alcune linee L21 provenienti dallOlanda e dalla Germania settentrionale avrebbero invaso la Scandinavia (a partire dal 1.700 a.C.) portando con loro la sottoclade dominante, R1b-S21/U106 (vedi più sotto). La grande diffusione di L21 in Norvegia e in Islanda è dovuta ai vichinghi norvegesi che colonizzarono parti della Scozia e dellIrlanda rendendo schiavi alcuni nativi celtici e portandoli indietro in Norvegia o nella loro nuovo colonia dIslanda. Oggi, circa il 20% di tutte le linee maschili islandesi sono R1b-L21 di origine scozzese o irlandese.
In Francia, R1b-L21 è presente soprattutto nella Bretagna storica (incluse Mayenne e Vendée) e nella Bassa Normandia. Questa regione è stata ripopolata grazie alla massiccia immigrazione dei Britanni insulari avvenuta nel V secolo d.C. a causa della pressione esercitata dagli invasori anglosassoni. In ogni caso, è possibile che L21 fosse presente in Armorica già dalletà del bronzo o da quella del ferro, poiché le tribù della Confederazione Armorica dellantica Gallia possedevano già unidentità distinta da quella degli altri Galli e mantennero un legame molto stretto con le isole Britanniche almeno a partire dall età del bronzo atlantico.
Distribuzione dellaplogruppo R1b-L21 (S145) in Europa
 in Europa")
Il ramo gallico e iberico (DF27/S250)
I primi rami protoceltici R1b che dallEuropa centrale raggiunsero la Francia e la penisola Iberica furono probabilmente L21 e DF27. Se L21 pare abbia preso la strada settentrionale verso le isole Britanniche attraversando il Belgio e la Francia settentrionale, dallaltro lato DF27 si sarebbe diffuso in tutto il resto della Francia e soprattutto nella sua parte meridionale.
Nella penisola Iberica letà del bronzo non cominciò almeno fino al 1.800 a.C., e inizialmente rimase ristretta alle culture di El Argar e di Los Millares nel sudest della Spagna. Alcuni siti sporadici comparvero verso il 1.700 a.C in Castiglia e in Estremadura, e intorno al 1.500 a.C in Portogallo meridionale. In questi siti della prima età del bronzo sono state ritrovate solo qualche daga e qualche ascia in metallo, e dunque queste non possono essere considerate propriamente società delletà del bronzo quanto piuttosto società delletà del rame con qualche utensile in bronzo occasionale (forse importato). Queste culture potrebbero aver avuto origine a partire da piccoli gruppi di avventurieri R1b in cerca di facili conquiste in regioni dEuropa che ancora non conoscevano il nuovo metallo. Gli invasori sarebbero quindi diventati lélite dominante, si sarebbero riprodotti con le donne locali e, in capo a qualche generazione, la loro lingua indoeuropea sarebbe stata dimenticata e assorbita dalla lingua indigena (=> vedi Come i Baschi divennero R1b).
Martiniano et al. (2017) ha sequenziato i genomi di vari scheletri provenienti dalla penisola Iberica occidentale e risalenti al neolitico medio o tardo, al calcolitico e alla media età del bronzo (la prima età del bronzo non raggiunse mai queste regioni). Ha così scoperto che gli individui neolitici e calcolitici appartenevano agli aplogruppi I*, I2a1 e G2a. Al contrario, tutti e tre i campioni portoghesi delletà del bronzo appartenevano a R1b (un M269 e due P312), nonostante possedessero linee materne iberiche tipiche del neolitico (H1, U5b3, X2b) e mancassero completamente di qualsiasi miscela ricollegabile alle steppe. Ciò concorda con lo scenario secondo cui gli uomini R1b indoeuropei sarebbero arrivati nella penisola Iberica in piccoli gruppi a partire dal 1.800 a.C., e avrebbero preso come mogli donne locali finendo col diluire il loro DNA a ogni nuova generazione fino a che, in qualche secolo, la miscela delle steppe sarebbe sparita. Fu a quel punto che gli Indoeuropei raggiunsero il Portogallo. Oggi, Spagnoli e Portoghesi possiedono circa il 25% di miscela delle steppe, il che significa che vi furono altre importanti migrazioni indoeuropee che ebbero luogo nella tarda età del bronzo e nelletà del ferro.
La penisola Iberica non entrò a pieno nelletà del bronzo almeno fino al XII secolo a.C., quando la cultura dei campi durne (1.300-1.200 a.C.) si espanse dalla Germania verso la Catalogna passando per la Francia meridionale. La successiva cultura di Hallstatt (1.200-750 a.C.) si diffuse in tutta a penisola (specialmente nella sua metà occidentale). Questo periodo fa parte della più ampia età del bronzo atlantico (1.300-700 a.C.), un periodo durante il quale la penisola Iberica era connessa al resto dellEuropa occidentale da una fitta rete di commerci.
È difficile dire quando esattamente DF27 arrivò nella penisola Iberica. Considerando la sua massiccia presenza in queste zone e in Francia sudoccidentale, è probabile che DF27 sia arrivato molto presto, in un periodo compreso tra il 1.800 e il 1.300 a.C. se non addirittura prima. Gli avventurieri R1b, infatti, potrebbero essersi infiltrati nella cultura del vaso campaniforme, come sembra abbiano fatto nel resto dellEuropa occidentale dal 2.300 al 1.800 a.C. Letà del bronzo atlantico potrebbe corrispondere al periodo in cui DF27 si distribuì capillarmente nella penisola Iberica per poi finire, seguendo le vie di commercio atlantiche, fin nelle isole Britanniche, in Olanda e in Scandinavia.
Distribuzione dellaplogruppo R1b-DF27 in Europa

Il ramo italo-celtico (S28/U152/PF6570)
A partire dal 1.300 a.C. circa, anche grazie allabbondanza di metalli fiorì intorno alle Alpi una nuova cultura delletà del bronzo. Questa gettò le basi della cultura classica dei Celti. In realtà diede origine a tre culture strettamente legate tra loro: la cultura dei campi durne e la cultura di Hallstatt (dal 1.200 a.C.) e quella di La Tène (dal 450 a.C.). Dopo lespansione della cultura di Unetice in Europa occidentale, tra il 2.300 e il 1.800 a.C., il periodo che comprende le culture del campo durne/Hallstatt/La Tène rappresenta la seconda maggior espansione di R1b dallEuropa centrale verso lAtlantico, la Scandinavia settentrionale, la valle del Danubio orientale, la Grecia, lAnatolia, lUcraina e la Russia, e forse fino al bacino del fiume Tarim in Cina nordoccidentale (=> vedi Mummie Tarim.
Letà del ferro celtica (tarda Hallstatt, dall800 a.C.) potrebbe essere cominciata grazie ai contatti, mai interrotti, con le steppe e con il Caucaso settentrionale, in particolare con la cultura di Koban (1.100-400 a.C.).
I Celti alpini della cultura di Hallstatt sono associati, seppure non in modo esclusivo, alla mutazione S28 (anche detta U152 o PF6570) R1b-S28 sarebbe sceso in Italia attraversando le Alpi in ondate successive a partire dal 1.700 a.C. È in questo periodo che nella valle del Po comparve la cultura Terramare Dal 1.200 a.C., varie tribù derivate dalla cultura di Hallstatt fondarono la cultura di Villanova (vedi sotto). Probabilmente fu proprio questa migrazione a portare le tribù italiche in Italia, le quali apparterrebbero soprattutto alla clade Z56 di R1b-S28. Durante letà del ferro, lespansione della cultura di La Tène in Svizzera è associata alla diffusione del ramo Z36, che avrebbe dato origine ai Belgi intorno al moderno Belgio e in Renania, ai Galli in Francia e ai Celti cisalpini in Italia.

Un tratto linguistico comune allitalico e alle lingue celtiche/galliche/britanniche, connesso con lespansione della cultura di Hallstatt, è la transizione dalloriginale suono indoeuropeo *Kw verso il suono *p. I linguisti classificano queste lingue appunto come celtico P, in opposizione al celtico Q. Si pensa che questa transizione sia avvenuta a causa dellincapacità di pronunciare il suono *Kw da parte delle popolazioni preindoeuropee dellEuropa centrale, della Gallia e dellItalia, che in origine parlavano dialetti afro-asiatici evolutisi dalle lingue del Vicino Oriente ed ereditate con la rivoluzione neolitica. Gli Etruschi, nonostante siano arrivati più tardi, forse dal Mediterraneo orientale, cadono anchessi in questa categoria. È stato recentemente dimostrato che le lingue celtiche hanno preso in prestito parte della loro grammatica dalle lingue afro-asiatiche. Questo passaggio potrebbe essere avvenuto quando i proto-celto-italici si spostarono dalle steppe al bacino del Danubio mischiandosi con le popolazioni di agricoltori originari del Vicino Oriente e appartenenti agli aplogruppi E1b1b, G2a, J e T. In ogni caso, una transizione così precoce non spiegherebbe perché il celtico Q e le lingue germaniche non siano andate incontro alla stessa mutazione. È dunque più plausibile che la transizione sia avvenuta dopo che i proto-italo-celti e i proto-germani si erano già espansi per la prima volta in Europa orientale e settentrionale. Il legame tra il celtico P e S26/U152 suggerisce che il passaggio sia avvenuto sulle Alpi dopo il 1.800 a.C., ma prima dellinvasione dItalia da parte delle tribù italiche (1.200 a.C. circa).
Lespansione in Italia della cultura dei campi durne e di quella di Hallstatt è evidente nella forma assunta dalla cultura Villanoviana (circa 1.100-700 a.C.), la quale presenta unimpressionante somiglianza con queste culture della Bavaria e dellUPPER AUSTRIA. La cultura di Villanova segna un chiaro punto di rottura con la precedente cultura di terramare. Nonostante entrambe praticassero lincinerazione, va ricordato che i Terramare riponevano i resti in ossari comuni, proprio come i loro predecessori neolitici del Vicino Oriente, mentre i Villanoviani utilizzavano sia urne biconiche tipiche della cultura dei campi durne che tombe contenenti gioielli, armature di bronzo
di cavalli per le élite. Ciò mostra per la pima volta in Italia lo sviluppo di quella società fortemente gerarchica tipica delle culture indoeuropee. Insieme alle sepolture, apparerò per la prima volta decorazioni tisicamente indoeuropee come per esempio le svastiche. Pur essendo in origine una cultura delletà del bronzo, i Villanoviani introdussero la metallurgia del ferro nella penisola Italica più o meno in contemporanea alla sua comparsa nella cultura di Hallstatt, rinforzando così lidea di un legame tra le due culture. Con tutta probabilità, la diffusione della cultura di Villanova rappresenta proprio la colonizzazione italica della penisola. La più alta percentuale di R1b-S28 si trova precisamente dove si insediarono i Villanoviani intorno alla moderna Toscana e allEmilia-Romagna. Alla cultura di Villanova succedette la civiltà Etrusca che mostra segni di continuità con la prima ma anche nuovi elementi ibridi originari dellAsia occidentale, probabilmente introdotti da coloni anatolici (che sarebbero appartenuti a un miscuglio di E1b1b, G2a, J1, J2 e R1b-L23).
Distribuzione dellaplogruppo R1b-S28 (U152) in Europa
 in Europa")
Il ramo germanico (S21/U106/M405)
Il principale ramo protogermanico della famiglia indoeuropea è R1b-S21 (U106 o M405). Questo aplogruppo si trova con alte concentrazioni in Olanda e in Germania nordoccidentale. È probabile che le linee R1b-S21 si siano espanse in questa regione grazie a un effetto del fondatore durante il periodo di Unetice, e che siano poi penetrate in Scandinavia intorno al 1.700 a.C. (probabilmente insieme a R1a-L664). Avrebbero così dato origine a un nuovo periodo, letà del bronzo nordica (1.700-500 a.C.). R1b-S21 si sarebbe poi mescolato per più di un millennio con le preesistenti popolazioni scandinave rappresentate dagli aplogruppi I1, I2-L801 e R1a-Z284. Allinizio delletà del ferro germanica, intorno al 500 a.C., le genti scandinave avevano sviluppato una cultura e un linguaggio definitivamente germanici, ma erano divise in varie tribù che presentavano al loro interno diversi livelli dei vari aplogruppi. R1b-S21 divenne il tipo dominante tra le tribù germaniche occidentali, ma rimase una minoranza rispetto a I1 e a R1a in quelle orientali e settentrionali, incluse quelle originarie della Svezia come i Goti, i Vandali e i Longobardi.
La presenza di R1b-S21 in altre parti dEuropa può essere attribuita quasi esclusivamente alle migrazioni germaniche che ebbero luogo intono al III e al X secolo d.C. I Frisoni e gli Anglosassoni disseminarono questo aplogruppo in Inghilterra e nelle pianure scozzesi; i Franchi lo fecero in Belgio e in Francia; i Burgundi in Francia orientale; gli Svevi in Galizia e nel nord del Portogallo; i Longobardi in Austria e in Italia. I Goti diedero il loro contributo alla propagazione di S21 in Europa orientale, ma pare che le loro linee germaniche siano andate diluendosi grazie allunione con le popolazioni slave e balcaniche, così che il loro impatto genetico sullItalia, sulla Francia e sulla Spagna non fu mai davvero significativo. Più tardi, i vichinghi danesi e norvegesi avrebbero a loro volta contribuito alla diffusione di R1b-S21 (insieme a I1, I2b1 e R1a) in quasi tutta lEuropa occidentale, ma soprattutto in Islanda, nelle isole Britanniche, in Normandia e in Italia meridionale.
A partire dal basso medioevo fino allinizio del XX secolo, i Germani si sono espansi in quasi tutta la Polonia spingendosi fino alla Lettonia a nordest e fino alla Romania a sudest. Durante questo periodo gli Austriaci hanno costruito un impero comprendente lodierna Repubblica Ceca, la Slovacchia, lUngheria, la Slovenia, la Croazia, la Serbia e parte della Romania, dellUcraina occidentale, e della Polonia meridionale. Diversi secoli di influenza tedesca e austriaca sullEuropa centrale e orientale appaiono evidenti nelle basse percentuali di linee germaniche che si trovano oggi tra queste popolazioni. In Romania il 4% della popolazione considera se stessa tedesca. La bassa incidenza di R1b-S21 in Finlandia, in Estonia e in Lettonia può essere attribuita alle dominazioni svedese e danese che cominciò nel basso medioevo e terminò sul finire del XIX secolo.
Distribuzione dellaplogruppo R1b-S21 (U106) in Europa
 in Europa")
| Come R1b rimpiazzò quasi tutte le linee più antiche dellEuropa occidentale |
|---|
|
Fino a poco tempo fa, a causa della sua forte presenza in Europa occidentale, si pensava che R1b avesse avuto origine proprio in questarea. La teoria voleva che R1b rappresentasse gli Europei paleolitici (Cro-Magnon) che durante lultimo massimo glaciale si erano rifugiati nelle regioni franco-cantabriche. Da qui, quando i ghiacci si erano ormai ritirati, avrebbero poi colonizzato nuovamente lEuropa centrale e settentrionale. Lanalisi filogenetica di R1b dimostra limpossibilità di questo scenario, poiché le più antiche cladi di R1b si trovano invariabilmente in Asia centrale e nel Medioriente, mentre quelle più recenti si trovano in Europa occidentale e settentrionale. Cè un chiaro gradiente che va da Est a Ovest seguendo la migrazione delle genti R1b (vedi la mappa più in alto). Il periodo in cui avvenne la migrazione principale dalle coste del mar Nero verso lEuropa centrale coincide con la finestra temporale durante la quale gli Indoeuropei avrebbero invaso lEuropa, e questa finestra è sovrapponibile allintroduzione del bronzo in Europa occidentale e alla proliferazione delle lingue italo-celtiche e germaniche.
A lungo gli storici e gli archeologi hanno dibattuto se le migrazioni indoeuropee siano state vere e proprie migrazioni di massa, o se piuttosto non siano state il frutto della diffusione culturale di una lingua e di diverse tecnologie, portate da un piccolo numero di invasori. È possibile che nessuna di queste due opzioni sia corretta. Chi propone la teoria della diffusione culturale cerca di convincerci che R1b sia nativo dellEuropa occidentale, e che solo R1a rappresenti gli Indoeuropei. Il problema è che laplogruppo R apparve in Asia centrale, e ancora oggi R2 è confinato in asia meridionale e centrale, e R1a e le sottocladi più antiche di R1b si trovano anchesse in Asia centrale. Letà delle sottocladi di R1b presenti in Europa coincide con letà del bronzo. Di conseguenza R1b deve aver rimpiazzato la maggior parte delle linee di DNA Y native dellEuropa a partire da questo periodo in poi.
In ogni caso, è risulta complicato anche abbracciare la teoria di una migrazione massiva che avrebbe annichilito quasi completamente la popolazione paleolitica. Effettivamente gli Europei occidentali moderni appaiono diversi in Irlanda, in Olanda, in Aquitania o in Portogallo, nonostante in tutte queste regioni domini R1b. Gli studi sul DNA autosomico hanno confermato che la popolazione dellEuropa occidentale è tuttaltro che omogenea. Anche alcune delle linee materne (DNA mt) sembrano avere origine paleolitica (H1, H3, U5, V). Quel che facilmente ci si dimentica è che, affinché delle linee paterne siano rimpiazzate rapidamente, non è necessario postulare un esodo di massa. Qui mostriamo il motivo.
- Poligamia. A differenza delle donne, non esiste un limite al numero di figli generabili da un uomo. Di solito gli uomini di potere hanno più figli rispetto ai loro subalterni. Ciò era particolarmente vero nelle società primitive dove la poligamia era spesso la norma per i capitribù e per i re.
- Status e potere. Equipaggiati con armi in bronzo e a dorso dei cavalli, gli Indoeuropei avrebbero soggiogato facilmente gli agricoltori neolitici e ancora più agilmente gli ultimi cacciatori-raccoglitori europei. Dove non sterminarono gli indigeni, i nuovi arrivati sarebbero diventati la classe dominante, con una moltitudine di capi locali, di re e di nobili (i Celti e i Germani delletà del bronzo vivevano in piccoli comunità raccolte in villaggi ognuno con un suo capo, e ogni parte della piccola tribù era guidata da un re) con probabilità di riprodursi superiori alla media.
- Sbilanciamento di genere. Le armate degli invasori di solito sono composte da una maggioranza di uomini. Gli uomini devono quindi trovare delle compagne tra le popolazioni conquistate. La guerra era un affare per lo più maschile, e i perdenti soffrivano perdite consistenti, lasciando così più donne disponibili per i vincitori.
- Politica militare aggressiva.. 4. Gli Indoeuropei erano guerrieri e seguivano un codice incentrato sul valore militare che enfatizzava atti di eroismo e di prodezza. La loro superiorità tecnologica (armi in metallo, veicoli dotati di ruote e trainati da cavalli) e la loro visione della vita avrebbero permesso loro di sconfiggere qualsiasi popolo incontrassero sul cammino che non avesse un esercito organizzato e dotato di armi in metallo (cioè chiunque a parte le civiltà mediorientali).
- Predisposizione genetica al concepimento di maschi. Il ruolo principale del cromosoma Y è la regolazione della produzione di sperma. I vari aplogruppi Y sono determinati dalle mutazioni che di volta in volta appaiono sul cromosoma Y. Ogni mutazione potrebbe in teoria influenzare la produzione di sperma e la sua motilità. Alcune ricerche preliminari hanno già stabilito un legame tra certi aplogruppi e un aumento o una diminuzione della motilità dello sperma. Più alta è la motilità, più è alta la probabilità di concepire un maschio. È del tutto possibile che R1b abbia determinato uno sbilanciamento di genere favorendo la generazione di un maggior numero di individui maschi. Anche solo un piccolo cambio percentuale contribuirebbe significativamente al rimpiazzo delle altre linee, con un effetto di accumulazione lungo i millenni. Non tutte le sottocladi di R1b avrebbero questo bias. Il bias esiste solo in relazione agli altri aplogruppi che si trovano nella stessa popolazione. È ben possibile che le più recenti cladi di R1b avessero un vantaggio significativo in confronto agli aplogruppi più vecchi come I2 e E-V13. Leggi altro
Secondo questo modello, la sostituzione delle linee paterne diventa rapidamente esponenziale. Per esempio: si immaginino cento Indoeuropei che conquistano una tribù di mille Europei indigeni (rapporto di 1:10). I morti in battaglia fanno sì che tra i conquistati ci sia una maggior numero di donne rispetto agli uomini. Diciamo che tra i sopravvissuti indigeni ci sono settecento donne e trecento uomini. Supponiamo poi che tra gli Indoeuropei vittoriosi letà adulta sia raggiunta dal doppio dei nuovi nati. Ci sono diverse ragioni per giustificare questultimo punto: i vincitori potrebbero aver preso più mogli, o potrebbero avere delle concubine, o addirittura potrebbero aver stuprato le donne indigene. Uno status più elevato garantirebbe maggiori ricchezze e dunque una migliore possibilità di nutrire la propria prole, aumentando così le loro possibilità di raggiungere letà adulta. Un rapporto di nascite 2:1 in favore degli uomini è in realtà una stima conservativa, visto che è più che probabile che per la sensibilità delle genti delletà del bronzo sarebbe stato normale uccidere quasi tutti gli uomini sconfitti e rapire le loro donne (come attestato anche nellAntico Testamento). In uno scenario di questo tipo, alle nuove linee di DNA Y basterebbe qualche generazione per diventare le linee dominanti. Per esempio, se la prima generazione di Indoeuropei generasse due figli adulti per ciascun uomo, contro un figlio adulto per indigeno, già alla seconda generazione il numero di linee paterne indoeuropee passerebbe a duecento individui, a quattrocento nella terza, a ottocento nella quarta e a milleseicento nella quinta e così via. Nello stesso lasso di tempo, le linee indigene rimarrebbero a trecento individui per ciascuna generazione.
Basandosi su uno scenario di questo genere, le linee R1b sarebbero state benissimo in grado di rimpiazzare rapidamente le linee locali. Anche se i conquistatori indoeuropei ebbero solo qualche figlio in più degli uomini locali, questi sarebbero stati più che sufficienti affinché le linee R1b potessero diventare, entro qualche secolo, le linee dominanti. La cultura celtica durò per oltre mille anni in Europa continentale, prima che la conquista romana mettesse la parola fine ai privilegi dei capi e dei nobili. È un tempo più che sufficiente per permettere alle linee R1b di diffondersi tra il 50 e l80% della popolazione.
Le frequenze odierne di R1b formano un gradiente che va dalla costa Atlantica (dove la percentuale più alta) allEuropa centrale e orientale (dove è più bassa), per poi risalire fino alla sua terra dorigine, lAnatolia. Questo gradiente è dovuto quasi certamente al fatto che lagricoltura si radicò maggiormente in Europa orientale e poi in quella centrale, e qui le società raggiunsero una densità di popolazione più elevata, lasciando gli invasori R1b in minoranza. Inoltre, altri Indoeuropei, per esempio appartenenti alla cultura della ceramica cordata (R1a), erano già avanzati dalla moderna Russia e dallUcraina fino alla Germania e alla Scandinavia. Sarebbe stato difficile per le genti R1b scontrarsi con i cugini R1a che possedevano tecnologie e culture simili alle loro. Gli R1b pre-celto-germanici sarebbero dunque stati forzati a stabilirsi a ovest, prima nellarco alpino e poi conquistando la poco popolosa Europa occidentale.
|
Il ramo balcanico e asiatico (Z2103)
Il ramo asiatico
Sei campioni provenienti dallestremità orientale dellorizzonte Jamna, nella regione Idel-Ural, analizzati da Haak et al. (2015) sono tutti risultati essere R1b. Quattro di questi sono positivi per la mutazione Z2103. Con tutta probabilità, R1b-Z2103 fu una delle linee principali della cultura di Polatvka, che succedette alla cultura Jamna tra il fiume Volga e i monti Urali. In seguito si fuse con la cultura di Abashevo (presumibilmente appartenente al ramo R1a-Z93) per poi formare la cultura Sintashta. Grazie a un effetto del fondatore, o attraverso la dominazione politica, le linee R1a-Z93 sarebbero riuscite a sopraffare R1b-Z2103 appena dopo la sua espansione in Asia centrale e meridionale, nonostante alcune importante sacche di Z2103 sopravvissero in Bashkorostan, in Turkmenistan e nellUiguristan (Turkestan cinese).
R1b-Z2103, proprio come R1a-Z93, sarebbe diventata una linea indoiraniana. Ciò è vero soprattutto per due sue sottocladi: L227.1 e L584. La prima si trova dalla Russia allAsia centrale e dallIndia al Medioriente, esattamente come la sottoclade Z93 di R1a-L657. L227.1 può essere associata alla cultura di Andronovo e al complesso archeologico battriano-margiano, così come alle migrazioni indoarie. R1b-L584 si trova soprattutto in Iran, nellIraq settentrionale, nel Caucaso meridionale e in Turchia, e correla soprattutto con il ramo iraniano degli Indoeuropei, che include i Persiani, i Curdi e gli Sciti.
Distribuzione dellaplogruppo R1b-ht35 (L23, L11, L51 & Z2103) in Europa
 in Europa")
Il ramo anatolico
Gli Ittiti (circa 2.000-1.178 a.C.) furono i primi Indoeuropei che riuscirono a sconfiggere le potenti armate mesopotamiche ed egiziane. Ci sono due ipotesi riguardo alla loro origine. Secondo la prima, gli Ittiti sarebbero arrivati dai Balcani orientali e avrebbero invaso lAnatolia attraversando il Bosforo. Ciò starebbe a significare che appartenevano alla sottoclade L23* o a Z2103. Nel secondo scenario, gli Ittiti sarebbero una ramificazione del tardo periodo della cultura Majkop, e avrebbero attraversato il Caucaso per poi conquistare i regni di Hatti (forse dopo essere stati scalzati dal Caucaso settentrionale dalle genti R1a della cultura delle catacombe). In questo caso sarebbero appartenuti alle sottocladi R1b-Z2103 o R1b-PF7562. La prima ipotesi ha il vantaggio di far partire da un unico nucleo, i Balcani, tutte le espansioni indoeuropee R1b successive alla cultura di Jamna. Lipotesi Majkop, daltro canto, spiegherebbe perché il ramo anatolico delle lingue indoeuropee (ittita, luvio, lidio, palaico) appaia così antico se comparato alle altre lingue indoeuropee, le quali avrebbero avuto tutte origine dalla cultura di Jamna e non da quella di Majkop.
Ci sono numerose prove archeologiche e linguistiche a favore di un legame tra Troia e le culture delle steppe R1b. Esse confermerebbero lorigine indoeuropea di questa città. I Troiani parlavano il Luvio, erano imparentati con gli Ittiti (quindi Indoeuropei), e avevano legami culturali con le culture delle steppe pontico-caspiche. La prima città di Troia risale al 3.000 a.C., nel bel mezzo del periodo Majkop. Troia potrebbe essere stata una colonia fondata dalle genti Majkop per assicurarsi il controllo delle vie commerciali tra il mar Nero e lEgeo. La fondazione di Troia coincide esattamente con il periodo durante il quale vennero costruite le prime galee. Se si considera lantichità della fondazione di questa città, è probabile che la sua linea paterna dominante fossero R1b-M269 o L23.
I Frigi e i Protoarmeni sono altre due tribù indoeuropee nate nei Balcani. Sembra che entrambe siano emigrate in Anatolia intorno al 1.200 a.C., durante il i grandi sconvolgimenti del Mediterraneo orientale (vedi sotto). I Frigi (o Brigi) fondarono un regno (1.200-700 a.C.) in Anatolia centroccidentale, prendendo il controllo dellagonizzante impero Ittita. Gli Armeni attraversarono tutta lAnatolia fino a lago di Van, e si fermarono sulle montagne armene. Oggi, il 30% degli Armeni appartiene allaplogruppo R1b. La gran parte di questi presenta la mutazione L584 di Z2103 (=> vedi Le migrazioni indoeuropee in Armenia).
La maggior parte degli R1b che si trovano oggi in Grecia fanno parte della varietà balcanica Z2103. Vi è anche una minoranza protoceltica S116/P312 e una celtica italo-alpina S28/U152. Z2103 potrebbe essere sceso dallAlbania o dalla Macedonia durante le invasioni dei Dori (vedi sotto), che si pensa siano avvenute nel XII secolo a.C. La lingua dei Dori sembra fosse sufficientemente simile al miceneo da permettere la mutua comprensione e la sua adozione da parte dei popoli locali. Anche i Micenei potrebbero aver portato un po di R1b (probabilmente Z2103), ma le loro origini possono essere fatte risalire, attraverso le prove archeologiche, alla cultura delle Catacombe e al fenomeno Seima-Turbino delle foreste-steppe settentrionali, che li renderebbe una tribù a maggioranza R1a.
LS116 e lS28 greci e anatolici potrebbero essere attribuiti alle invasioni celtiche del periodo di La Tène avvenute nel III secolo d.C. Certo, anche i Romani portarono linee S28 (=> vedi Genetica degli Italiani), così come più tardi i Veneziani, specie nelle isole. Le cladi più antiche di R1b, come P25 e V88, rappresentano solo una piccola minoranza e sarebbero arrivate insieme a E1b1b, G2a e J2 dal Medioriente.
| I grandi sconvolgimenti del 1200 a.C. |
|---|
|
Il 1.200 a.C. rappresenta un punto di svolta nella storia europea e del Medioriente. In Europa centrale, la cultura dei campi durne si evolvette nella cultura di Hallstatt, tradizionalmente associata alla civiltà classica dei Celti e che ebbe uninfluenza fondamentale sullo sviluppo dellantica Roma. Nelle steppe pontiche, la cultura di Srubna fece posto ai Cimmeri, genti nomadi che parlavano una lingua iraniana o tracica. La cultura colchica (1.200-600 a.C.) delletà del ferro nacque nella regione del Caucaso settentrionale. La sua successiva espansione nelle regioni meridionali corrisponde alla prima menzione storica del ramo protoarmeno delle lingue indoeuropee (circa 1.200 a.C.). Nel Levante centrale, i Fenici cominciarono ad affermarsi come una significativa potenza marittima e iniziarono a costruire il loro impero commerciale intorno al Mediterraneo meridionale.
Tuttavia, levento più importante di questo periodo è sicuramente la distruzione delle civiltà mediorientali avvenuta forse a opera dei Popoli del Mare. La grande catastrofe che coinvolse linterno bacino mediterraneo orientale, dalla Grecia allEgitto intorno al 1.200 a.C., è un soggetto ancora oggi molto controverso. Lidentità dei Popoli del Mare è stata oggetto di numerose speculazioni. Quel che è certo, è che tutte le società palaziali del Medioriente andarono incontro a terribili atti di distruzione, saccheggi e razzie. La spiegazione più diffusa è linvasione da parte di popolazioni guerriere provenienti dal Nord e tecnologicamente più avanzate. Questi potrebbero esser stati gli Indoeuropei scesi dalle steppe attraverso i Balcani, oppure genti caucasiche (G2a, J1, J2a, T1a) legate allespansione della prima cultura di Kura-Araxes in Anatolia orientale e nel Levante.
La capitale Ittita, Hattusa, venne distrutta nel 1.200 a.C. e, verso il 1.160 a.C., limpero collassò, probabilmente sotto la pressione dei Frigi e degli Armeni arrivati dai Balcani. Le città micenee furono devastate e abbandonante per tutto il XII secolo d.C., fino al totale collasso della civiltà micenea avvenuto nel 1.100 a.C. Il regno di Ugarit in Siria, fu annichilito e la sua capitale non venne mai più ricostruita. Altre città del Levante, di Cipro e di Creta furono bruciate e lasciate in totale abbandono per diverse generazioni. Gli Egiziani dovettero respingere gli assalti dei Filistei dallest e dei Libici dallovest, entrambe popolazioni di probabile origine indoeuropea. I libici erano accompagnati da mercenari originari del nord (gli Ekwesh, i Teresh, i Lukka, gli Sherden e gli Shekelesh), le cui origini sono incerte e sono state individuate in Anatolia, in Grecia e/o in Italia meridionale.
La devastazione della Grecia seguì di poco la leggendaria Trojan War (1.194-1.187 a.C.). È stato proposto che i Dori, una popolazione indoeuropea proveniente dai Balcani (forse dalla moderna Bulgaria o dalla Macedonia), riuscirono a invadere la Grecia micenea perché indebolita dalle guerre troiane, e poi si stabilirono proprio in questa regione come uno dei tre maggiori gruppi etnici. Le regioni doriche della Grecia classica, dove si parlavano i dialetti dorici, sono essenzialmente il Peloponneso meridionale e orientale, Creta e Rodi, e sono anche le aree della Grecia con la più alta percentuale di R1b-Z2103.
Unaltra teoria vuole che gli Illiri, emigrati dallEuropa nordorientale verso i Balcani, abbiano scacciato le tribù indoeuropee che già abitavano quelle terre. I Dori allora avrebbero dovuto spostarsi in Grecia, i Frigi in Anatolia nordoccidentale e i Libu, dopo un fallito tentativo di conquista del delta del Nilo in Egitto, in Libia. I Filistei, arrivati forse dallAnatolia, si stabilirono definitivamente in Palestina intorno al 1.200 a.C., impossibilitati a entrare in Egitto.
|
Altre migrazioni R1b
Durante lantichità e durante il medioevo ci furono altre migrazioni che, a partire dallEuropa, coinvolsero il Vicino Oriente e lAsia centrale. R1b-S28 (U152) è stato trovato in Romania, in Turchia, in Baschiria settentrionale (unimpressionante 71.5% della popolazione locale secondo Myres et al.), e al confine tra il Kazakistan e il Kyrgyzstan. In parte fu sicuramente portato dai Celti di La Tène, noti per essersi avventurati lungo le rive del Danubio e per aver creato il regno di Galazia in Anatolia centrale. Il resto potrebbe benissimo avere origine romana, dato che R1b-S28 è la forma dominante di R1b nella penisola Italiana. Qualcuno ha suggerito che alcune legioni romane perdute si siano spinte fino allAsia centrale o fino alla Cina e abbiano preso in moglie donne locali lasciando la loro impronta genetica in Asia almeno in alcune sacche isolate. Una versione meno romanzesca vuole che allorigine di queste sacche ci siano invece i mercanti romani che, attraverso la via della seta nata verso il II secolo d.C., si stabilirono in Cina.
Una piccola percentuale delle sottocladi di R1b dellEuropa occidentale sono state trovate anche tra le comunità cristiane del Libano. Questi rappresentano molto probabilmente i discendenti dei Crociati.
La persistenza di lattasi e i mandriani R1b
Il lattosio (zucchero del latte) è una componente essenziale del latte materno ed è consumato dai neonati. La sua digestione è possibile grazie a un enzima, chiamato lattasi, che spezza il lattosio in zuccheri più semplici che possono poi essere assorbiti dalle pareti intestinali e raggiungere il flusso sanguigno. Nella maggior parte dei mammiferi (umani inclusi), la produzione dellenzima è drasticamente ridotta poco dopo lo svezzamento. Come risultato, gli adolescenti e gli adulti diventano intolleranti al lattosio. Ciò è vero per una buona parte della popolazione mondiale. Alcune persone possiedono una mutazione genetica che permette la produzione di lattasi anche durante letà adulta. Questo fenomeno si chiama persistenza di lattasi (LP). Essa è particolarmente diffusa tra gli Europei nordoccidentali discendenti delle antiche genti celtiche e germaniche e in alcune parti dellAfrica dove lallevamento dei bovini è stato praticato per migliaia di anni. La più alta incidenza degli alleli per la persistenza di lattasi, noti ai genetisti come -13.910*T (rs4988235) e -22018*A (rs182549), si trova in Scandinavia, in Olanda, in Gran Bretagna, in Irlanda e tra i Baschi. Le popolazioni subsahariane che presentano la persistenza di lattasi presentano mutazioni diverse, come per esempio -14010*C, -13915*G e -13907*G.
Gli uomini R1b sono considerati i primi ad aver addomesticato con successo i bovini e i primi, durante il neolitico preceramico, ad aver sviluppato uno stile di vita basato sullallevamento delle mandrie (vedi la sezione sul neolitico). In cerca di pascoli per le loro mucche, le tribù R1b migrarono dal Medioriente verso le savane dellAfrica settentrionale (che è poi andata incontro alla desertificazione diventando il Sahara) e verso le steppe pontiche della Russia meridionale e dellUcraina. Per diversi millenni nessunaltra popolazione fu dipendente dalle mandrie bovine quanto queste tribù R1b.
È noto che la maggior parte delle società neolitiche dipendenti dai bovini consumava almeno in parte il latte animale e lo utilizzava per fare il formaggio (il formaggio contiene meno lattosio ed è più facile da digerire per chi è intollerante). Nella gran parte dEuropa, nel Medioriente e in Asia meridionale, lallevamento animale riguardava soprattutto ovini e caprini, più adatti agli ambienti montagnosi del bacino mediterraneo, dellAnatolia e dellIran. Questi animali potevano facilmente essere protetti dagli agricoltori di cereali nei loro villaggi, mentre i bovini avevano bisogno di ampi prati per pascolare che in Medioriente scarseggiavano. A volte alcuni bovini domestici erano presenti, in piccolo numero, anche tra le popolazioni neolitiche, ma le società che basavano su di esse la propria economia erano le tribù R1b delle steppe pontiche e del nord africa. Ancora oggi, i mandriani del Sahel come i Fulani e gli Hausa, che discendono dai migranti neolitici R1b-V88 mediorientali, mantengono le loro mandrie di bovini. È tra questi mandriani che la pressione selettiva per la persistenza di lattasi sarebbe stata più forte.
Sono state avanzate diverse ipotesi dai genetisti e dai biologi dellevoluzione riguardo alla persistenza di lattasi. Sono stati analizzati più di cento genomi antichi risalenti al mesolitico, al neolitico e alletà del bronzo europea e della Siria; lallele -13910*T è stato identificato solo negli individui del tardo neolitico/calcolitico e delletà del bronzo. Lorigine della mutazione non è il punto fondamentale della discussione, dato che essa potrebbe essere stata presente nel pool genetico umano, con una bassa incidenza, per decine di migliaia di anni prima che fosse selezionata positivamente nelle società di mandriani. Quel che è certo è che gli individui delletà del bronzo associati allarrivo degli Indoeuropei dalle steppe pontiche possedevano già percentuali relativamente elevate dellallele LP. Per esempio, lo si può ritrovare con una frequenza del 27% (vedi (see Schilz 2006) tra 13 individui provenienti dalla grotta Lichtenstein in Germania, i quali appartenevano alla cultura dei campi durne ed erano un mix di aplogruppi Y R1b, R1a e I2a2b.
Oggi lallele LP è proporzionale alla percentuale di R1b e, in misura minore, alla percentuale di R1a che si trova in una data popolazione. Nelle isole Britanniche, nei Paesi Bassi e nella Scandinavia sudoccidentale, dove LP raggiunge le frequenze massime a livello mondiale, la somma di R1a e R1b eccede il 70% della popolazione. Nella penisola Iberica la percentuale più alta si trova tra i Baschi che presentano anche lincidenza massima di R1b. In Italia, LP è diffuso soprattutto al nord, proprio come R1b. Le percentuali più basse di LP in Europa si trovano in Italia meridionale, in Grecia e nei Balcani, cioè le regioni con il minor numero di linee R1b.
Tishkoff et al. (2017) ha confermato che le genti Hausa e Fulani, due tribù del Sahel con unalta incidenza di R1b-V88, possiedono lo stesso allele LP degli Europei, mentre i pastori dellAfrica orientale con unalta prevalenza di persistenza di lattasi possiedono una mutazione completamente diversa e che sarebbe emersa indipendentemente. Questi risultati sono una prova che lallele -13,910*T abbia avuto origine con i primi mandriani R1b del Medioriente, i quali rappresentano i primi antenati sia degli Indoeuropei che delle tribù R1b-V88.
I popoli R1b diffusero i geni per la pelle e i capelli chiari o rossi
Ci sono oggi diverse prove della diffusione della mutazione A111T di SLC24A5, da parte dei popoli R1b e R1a. Questa mutazione spiega circa il 35% della differenza nella pigmentazione della pelle tra gli Europei e gli Africani e buona parte della sua variabilità in Asia meridionale. Il pattern di distribuzione dellallele A111T (rs1426654) ricalca quasi perfettamente la diffusione delle linee indoeuropee R1a e R1b in Europa, nel Medioriente, in Asia centrale e in Asia meridionale. È probabile che la mutazione sia passata alle altre popolazioni del Vicino Oriente allinizio del neolitico, cosa che spiegherebbe perché gli agricoltori neolitici europei avessero già lallele A111T (vedi Keller 2012 p.4, Lazaridis 2014 suppl. 7), anche se con frequenze più basse rispetto ai moderni Europei e agli Asiatici dellAsia centromeridionale.
Lallele per la pelle chiara si trova con una percentuale del 15-30% anche in diversi gruppi etnici nella parte settentrionale dellAfrica subsahariana, soprattutto nel Sahel e nelle savane abitate dalle tribù di mandriani R1b-V88 come i Fulani e gli Hausa. Ciò lascerebbe presupporre che lallele A111T fosse presente tra le tribù R1b già prima che questo aplogruppo si dividesse, durante il neolitico preceramico, in V88 e P297. Lincidenza di questo allele è la stessa nelle popolazioni R1a e nelle popolazioni R1b. Daltro canto, la mutazione A111T non è presente nel campione R* ritrovato in Siberia e risalente a 24.000 anni fa, e manca anche nella gran parte delle popolazioni R2 dellIndia sudorientale e dellAsia sudorientale in generale. Di conseguenza si può assumere che la mutazione sia apparsa nella linea R1* durante il paleolitico superiore, probabilmente in un periodo compreso tra i 20.000 e i 13.000 anni fa.
I capelli chiari sono un altro tratto associato agli Indoeuropei. I geni per gli occhi azzurri, invece, erano già presenti tra gli Europei mesolitici appartenenti allaplogruppo I. I geni per i capelli biondi correlano più che altro con la distribuzione dellaplogruppo R1a, ma quelli per i capelli rossi non sono mai stati trovati in Europa prima delletà del bronzo e pare siano stati diffusi soprattutto dagli R1b (=> vedi Lorigine dei capelli rossi).
Le linee materne (mtDNA) corrispondenti allaplogruppo R1b
Le linee discese dal ramo indoeuropeo di R1b (come L23) si trovano oggi in quasi tutta lEuropa e nelle vaste pianure del Medioriente e dellAsia centrale. Avanzando dalle steppe pontico caspiche, gli Indoeuropei R1b presero in moglie sia donne indoeuropee che donne locali. È dunque difficile scoprire quali fossero le linee di mtDNA originarie delle steppe o, ancora prima, del Medioriente neolitico o dellEurasia paleolitica. Ci sono due metodi per determinare quali fossero gli aplogruppi originari. Il primo consiste nellanalisi del DNA antico. Il secondo dipende dallanalisi delle linee materne che si trovano oggi nei luoghi dove si insediarono gli Indoeuropei, dallEuropa fino allAsia meridionale passando per lAsia centrale.
Antichi DNA mitocondriali collegati alle tribu R1b
Oggi sono stati testati centinaia di campioni di DNA mitocondriale risalenti al Neolitico, al mesolitico e perfino al paleolitico europeo. Questi studi hanno rivelato che laplogrupo R1b era ampiamente diffuso nellEuropa sud e nordorientale del mesolitico. Alcune tribù di cacciatori raccoglitori tra cui dominava R1b sono state trovate in Lettonia, intorno alle cosiddette Porte di ferro sul Danubio tra la Serbia e la Romania, e anche in Ucraina (sia del neolitico che del mesolitico) e nella regione del medio Volga in Russia. Le linee materne di questi popoli erano tipiche linee mesolitiche europee. Come altri gruppi appartenenti ad altre linee Y (C1a2, I*, I2a, I2c, R1a) queste tribù R1b appartenevano principalmente agli aplogruppi mitocondriali U5a e U5b, con una minoranza di U2 e U4. Alcuni R1b provenienti dalle Porte di ferro possedevano anche dei DNA mitocondriali meno tipici come K1, K1a, H13 e H40. In ogni caso, nessuno di questi individui mesolitici R1b è lantenato dei moderni Europei R1b, visto che nessuno possiede le mutazioni M269 o L23.
R1b-M269 si è evoluto in modo isolato per migliaia di anni, presumibilmente nellarea tra lAnatolia orientale e lIran nordoccidentale, per poi tornare in Europa orientale portandosi dietro la nuova mutazione L23 durante il calcolitico. Oggi, praticamente tutti gli uomini M269 privi della mutazione L23 si trovano in Armenia e lungo il bordo meridionale del mar Caspio in Azerbaijan e in Iran settentrionale. R1b-L23 ha portato la metallurgia nelle steppe pontiche caspiche ed è diventato la linea dellélite della nascente società indoeuropea, rimpiazzando rapidamente una buona parte delle altre linee delle steppe (I2a, J2b2a, Q1a, R1a, R1b) probabilmente anche grazie alla poligamia.
Molte linee materne della cultura di Jamna sono infatti diverse dalle precedenti linee mesolitiche e neolitiche delle steppe, e presentano una chiara origine in Asia occidentale e nel Caucaso. Queste nuove linee femminili includono H2b, H6a1, H15, K1b2, I1a, J2b, T1a, T2c1, W3, W6 and X2h. Come ci si aspetterebbe, molte di queste linee si trovano anche nelle altre culture delletà del bronzo associate alle lingue protoindoeuropee, incluse la cultura della ceramica cordata e di Unetice. Tra 46 campioni di DNA mitocondriale Jamna analizzati fino a giugno 2017, le linee mesolitiche U2, U4 e U5 rappresentano il 28% del totale. È possibile che alcuni campioni H e K abbiano avuto origine nelle steppe del mesolitico o per lo meno del neolitico.
Lidentificazione delle linee mitocondriali diffuse dalle migrazioni indosueorpee
Uno dei limiti del primo metodo è che necessita di un gran numero di campioni di DNA antico disponibili per lanalisi. Un altro problema è che funziona bene solo nellidentificazione degli aplogruppi portati dagli Indoeuropei durante letà del bronzo, ma non distingue quelli in comune con gli Europei mesolitici e neolitici. Per esempio, è certo che gli Indoeuropei delletà del bronzo condividessero con i cacciatori raccoglitori mesolitici gli aplogruppi U2, U4 e U5, e pare che anche alcune sottocladi di H e K fossero già presenti tra gli Europei mesolitici, incluse H1, H4, H10 H11, K1a e K1c. Se queste linee erano presenti nelle steppe pontico caspiche prima che R1b-L23 ebbe attraversato il Caucaso, sarebbero semplicemente state assimilate dai nuovi arrivati, ma potrebbero non aver niente a che fare con le genti originarie L23. Il secondo metodo cerca di rimediare a questo problema analizzando le linee mitocondriali degli Europei delletà del bronzo e i campioni moderni provenienti da quelle aree dellAsia in cui si stabilirono gli Indoeuropei.
Gli insediamenti isolati che si trovano in Asia, così come quelli del bacino del Tarim nella Cina nordoccidentale, della regione Altaj in Siberia meridionale, o della Battria e della Margiana in Asia centromeridionale, forniscono unopportunità unica per permetterci di trovare alcune linee mitocondriali europee in regioni che sono, dal punto di vista genetico, tipicamente asiatiche. Siccome in queste aree non ci fu nessunaltra importante insediamento storico europeo, il DNA mitocondriale europeo trovato in queste regioni dovrebbe necessariamente essere correlato a quello degli Indoeuropei delletà del bronzo e del ferro, come per esempio i portatori degli aplogruppi R1a e R1b. Lunica interferenza in questo caso sarebbe rappresentata dal DNA mitocondriale mediorientale, specialmente in quelle parti dellAsia centrale che sono a maggioranza musulmana come lo Xingjiang. Per fortuna, abbiamo unidea di quali aplogruppi-mt le genti E1b1b, J1, J2a e T1 avrebbero portato con loro. Essi sarebbero gli aplogruppi HV, N1, J, K, T2 e U3. In particolare pare che gli Sciti si siano ampiamente mescolati con i discendenti delle culture neolitiche dellAsia centromeridionale, visto che il DNA mitocondriale degli antichi Sciti contiene molti aplogruppi tipici di queste culture(insieme a H2a1, U2, U5 e le linee asiatiche orientali). Eppure, tutto il DNA-Y scita analizzato finora si è rivelato essere R1a, nonostante tra loro vi erano sicuramente delle linee R1b, J1 e J2.
Le linee europee o mediorientali trovate in Asia centrale e settentrionale includono HV, H2a1, H2b, H5a, H6, H7, H8, H15, I, J1b1, J1c (incluso J1c2m), K, N1a, R, T1, T2, U1, U2e, U3, U4, U5, U8, V, W, and X2. Se escludiamo gli aplogruppi degli agricoltori neolitici, cioè quelle più diffuse nei moderni paesi a maggioranza R1a, e se si escludono le line mediorientali più rare in Europa, ci rimangono come potenziali linee mitocondriali degli uomini R1b delletà del bronzo H2b, H5a, H6, H8c, H15, I, J1b1a, K1c1, K2b, T1a1, T2, U2e, U4, U5, U8, V7 and W. È probabile che H4, H5a, H6, K2b, U2, U4, U5 e U8 siano arrivate dalle popolazioni indigene delle steppe mesolitiche e neolitiche. Laplogruppo H5a è stato trovato nella cultura di Cucuteni-Trypillian e molto probabilmente entrò nel pool genetico delle steppe grazie alla pratica dei matrimoni misti.
Quali erano le originarie linee mitocondriali delle tribù calcolitiche R1b della Transcaucasia?
I candidati principali per le linee mitocondriali originarie associate alle tribù R1b-M269 e a R1b-L23 della Transcaucasia sono H8c, H15, I1a1, I2, I3, I4, J1b1a, T1a1, T2b (almeno alcune cladi come T2b2 e T2b4), V15 and W. A parte alcune cladi di T2b e W1, nessuno di questi aplogruppi è stato trovato né in Europa né tra i mediorientali neolitici o calcolitici prima delle migrazioni indoeuropee. Oggi J1b1a e T1a1 mostrano una correlazione particolarmente forte con la distribuzione dellaplogruppo Y R1b.
Gli aplogruppi V7 e T2b sono stati entrambi identificati tra alcuni reperti del periodo Majkop H2a1, I, K1c1, R1a e W sono molto comuni nel Caucaso settentrionale moderno. Alcune di queste linee (probabilmente H2a1, K1c1 e R1a, che oggi sono rare o assenti in Asia occidentale) potrebbero essere autoctone e potrebbero essere state assimilate solo in seguito, durante il periodo Majkop, dai migranti R1b-L23, mentre altre (gli aplogruppi I, T2b, V7 e W, oggi abbastanza comuni in Azerbaijan e in Iran nordoccidentale) potrebbero essere state portate proprio dagli R1b-L23 provenienti dallaltro lato del Caucaso.
Laplogruppo H2b si trova in Europa nordoccidentale, in Turchia, in Siberia, in Pakistan e in India. H8c si trova in Europa centrale e occidentale, in Georgia, in Armenia, in Asia centrale e negli Altaj. La loro bassa frequenza in Medioriente suggerisce unorigine caucasica, probabilmente nella sua parte settentrionale almeno per quanto riguarda H2b (come per H2a1).
Oggi H2a1 e V7a si trovano soprattutto nei paesi slavi settentrionali, ma potrebbero essere stati portati dalla minoranza R1b-L23 che si fuse con le tribù R1a della cultura della ceramica cordata (antenata delle genti slave). H2a1 infatti è stato trovato nella cultura della ceramica cordata e in quella di Unetice. Lo stesso sarebbe successo ad alcune cladi dellaplogruppo W. Oggi nei paesi a dominanza R1b si trovano specialmente W5 e W7, mentre altre cladi si trovano più diffuse in paesi a maggioranza R1a o presentano una distribuzione irregolare.
Laplogruppo I ha unorigine pan-caucasica e alcune delle sue cladi (I5, I6, I7) sono legate anche alla diffusione della cultura di Kura-Araxes. Quelli che correlano con più forza con il ramo indoeuropeo di R1b sono I1a1, I2, I3 and I4a.
Gli aplogruppi H15 e J1b1a si trovano entrambi in Armenia e in Iran. Solo la sottoclade J1b1a sembra correlata alla propagazione di R1b-L23. Altre sottocladi di J1b sono geograficamente limitate al Medioriente, in particolare alla regione tra il Caucaso e la penisola Arabica. J1b1a è stato ritrovato nelle culture della ceramica cordata di Unetice e nelletà del bronzo baltica.
T1a1 si trova in Transcaucasia, ma anche in Kurdistan e in Mesopotamia. Ciò indica una possibile connessione con lespansione di Uruk. T1a1 è stato trovato in molte culture preindoeuropee, incluse quella di Jamna, nella cultura della ceramica cordata, di Popapovka, di Srubnaya e in quelle deletà del bronzo baltica. Eppure, contrariamente ad altre linee di T1, non è mai stata trovata in campioni mesolitici o neolitici del Medioriente o dellEuropa, tranne che per un individuo ritrovato a Baalberge nella Germania del tardo neolitico (2.500 a.C.) contemporaneo allarrivo delle genti delle steppe in quella regione (dunque potrebbe essere semplicemente un migrante erroneamente identificato).
Laplogruppo V15 oggi si trova in Armenia e in Europa nordoccidentale ed è anchesso un buon candidato come linea originaria portata da R1b-L23 dalla Transcaucasia.
Quali erano le originarie linee mitocondriali delle tribù neolitiche R1b nel Vicino Oriente?
Si pensa che le tribù R1b abbiano addomesticato i bovini in questa regione verso i 10.500 anni fa, eppure si sarebbero spostati attraverso il Caucaso solo tra i 7.500 e i 6.500 anni fa. Per tre o quattro millenni, gli allevatori seminomadi R1b furono forzati a mescolarsi con i loro vicini mediorientali o caucasici. Un modo per determinare quali DNA mitocondriali portassero con loro queste tribù allinizio del neolitico consiste nel comparare questi aplogruppi con quelli dei gruppi etnici africani oggi noti per lalta incidenza di R1b-V88. Il gruppo più studiato è quello dei Fulani, il cui DNA mitocondriale include tre aplogruppi simil-europei H, J1b1a, U5 and V che tutti insieme formano il 15% delle loro linee materne. Questi aplogruppi sono stati identificati in tutti e quattro i paesi centrafricani analizzati, confermando la forte correlazione con laplogruppo R1b. In ogni caso, i loro H, V e U5 potrebbero essere arrivati dai Berberi dellAfrica nordoccidentale. Anche i Berberi possiedono R1b-V88, ma è possibile che una parte di esso sia arrivato attraverso altre migrazioni neolitiche, inclusa una seconda espansione a partire dalla penisola Iberica. I Berberi, infatti, portano le linee H1, H3, V1a1a, V5 e U5b1b1, tutte linee presenti anche nella penisola Iberica. U5b1b1 deriva dagli Europei occidentali mesolitici, ma al momento non è ancora chiaro come gli altri aplogruppi abbiano raggiunto la penisola Iberica o lAfrica nordoccidentale. Unipotesi è che sia arrivato dal Vicino Oriente durante il neolitico, forse insieme alle tribù R1b-V88.
Laplogruppo africano R1b-V88 e quello eurasiatico R1b-P297 si sarebbero separati circa 10.000 anni fa, quasi certamente in Europa orientale, dove portarono con loro soprattutto laplogruppo mitocondriale U5. Verso la fine dellultima glaciazione, alcuni uomini R1b sarebbero migrati dallEuropa orientale verso la regione del moderno Kurdistan accompagnati da donne appartenenti a U5. Appena dopo il loro arrivo, J1b1a (e forse V) sarebbe stata la prima linea indigena del Vicino Oriente ad essere assimilata dalle tribù R1b. R1b-V88 potrebbe aver assimilato donne H1 e H3 nel Levante prima di muoversi in Africa settentrionale, ma questa rimane una speculazione.
Individui famosi
Ramo R1b-S1200

L'Adams Surname Y-DNA Project ha comparato il DNA Y di 15 famiglie che presentano una storia legata a quella dei Presidenti degli Stati Uniti John Adams (secondo Presidente degli Stati Uniti) e di suo figlio John Quincy Adams (sesto Presidente). Tutti gli uomini analizzati condividono lo stesso aplogruppo che cade nella sottoclade S6849 (FGC2389) a valle di R1b-S1200
Ramo R1b-U106 (germanico)

Larmuseau et al. (2013) ha analizzato il DNA-Y di tre membri viventi della casa dei Borbone, uno discendente da Luigi XIII di Francia attraverso il Re Luigi Filippo I, e due da Luigi XIV attraverso Filippo V di Spagna. È stato concluso che tutti e tre questi uomini condividono lo stesso aplogruppo STR e appartengono allaplogruppo R1b-U106 (sottoclade Z381*). Questi risultati contraddicono gli studi più vecchi di Laluez-Fo et al. (2010), che aveva analizzato il DNA recuperato da un fazzoletto imbevuto nel presunto sangue di Luigi XVI dopo la sua esecuzione, e gli studi di Charlier et al. (2012), che aveva analizzato il DNA recuperato da una testa mozzata che sarebbe appartenuta a Enrico IV di Francia. Entrambi questi studi avevano identificato i resti come appartenenti allaplogruppo G2a. Tutti i Re di Francia discendono in linea paterna da Roberto il Forte (820-866), e a meno che qualche evento di non-paternità sia avvenuto prima di Luigi XIII, si può assumere che tutti i Re di Francia siano appartenuti alla stessa linea R1b-Z381. La casa dei Borboni comprende anche tutti i Re di Spagna da Filippo V (1683-1746) fino a oggi con il Re Juan Carlos, tutti i Re delle due Sicilie, i grandi duchi del Lussemburgo dal 1964, e ovviamente tutti i duchi di Orléans e i duchi di Borbone.

Grazie allanalisi di due discendenti noti della linea Wettin (analizzati da Brad Michael Little), la linea della casata di Wettin è stata identificata essere R1b-U106 (ramo Z381 > Z156 > Z305 > DF98+) I risultati hanno confermato che i due uomini hanno un antenato comune in Brad Michael Little. The results confirmed that the two men, who share a common ancestor in Francis, Duca di Saxe-Coburg-Saalfeld, e possiedono lo stesso aplogruppo e lo stesso aplotipo. I membri della casa di Wettin includono i Re Edoardo VII, Giorgio V, Edoardo VIII e Giorgio VI dInghilterra, tutti i Re del Belgio, i Re del Portogallo dal 1853 al 1910, i Re di Bulgaria dal 1887 al 1946, diversi Re di Polonia e gran duchi di Lituania, i margravi di Meissen dal 1075 al 1423, gli Elettori di Sassonia dal 1423 al 1806, i Re di Sassonia dal 1806 al 1918 e i signori di diversi ducati sassoni.

James K. Polk (1795-1849), lundicesimo Presidente degli Stati Uniti, era membro della sottoclade L48 almeno secondo i risultati del Polk-Pollock DNA Project. Era un discendente di William Polk/Pollock (1700-1757).

Secondo il Grant DNA Project, Ulysses S. Grant (1822-1885), (1822-1885), il diciottesimo Presidente degli Stati Uniti e comandante militare della guerra civile americana, apparteneva alla sottoclade di R1b-U106 FGC8590, a valle di L47 e Z159 (discendente di Matthew Graunt).

I primi due esseri umani il cui genoma è stato completamente sequenziato, James D. Watson, il co-scopritore della struttura del DNA, e il biologo Craig Venter sono entrambi parte dellaplogruppo R1b-S21 (U106), nonostante appartengano a sottocladi diverse (Z30, a valle di Z9 per Watson, e L45, a vale di L47 per Venter). Nonotante siano entrambi Americani, le loro rispettive linee paterne son scozzese e tedesca.
Altri membri famosi dellaplogruppo R1b-U106
- Malcolm Gladwell (nato nel 1963): è un giornalista inglese naturalizzato canadese, scrittore e speaker. Ha fatto parte degli autori del The New Yorker dal 1996. Ha scritto cinque libri, tutti apparsi nella lista dei Best Seller del The New Yorker. Il suo aplogruppo è stato rivelato da Faces of America.
Ramo R1b-L21 (celtico atlantico)

Il Buchanan DNA Project ha confermato che il quindicesimo Presidente degli Stati Uniti James Buchanan (1791-1868) discende dal clan scozzese dei Buchanan e dunque apparitene alla sottoclade S695 di R1b-L21, a valle di L1355 e S744.

William Ewart Gladstone (1809-1898), (1809-1898), un politico liberale britannico che sotto la Regina Vittoria fu Primo Ministro per la bellezza di quattro volte (un record), apparteneva a R1b-L21, almeno secondo il confronto della sua genealogia con i risultati del Gladstone DNA Project.

Lanalisi del DNA dei suoi discendenti ha confermato che Joseph Smith (1805-1844), il fondatore dei Mormoni e del Movimento dei Santi degli ultimi giorni, appartenenva allaplogruppo R1b-M222

Le analisi forensi sullo scheletro di Che Guevara (1928-1967), il rivoluzionario marxista argentino e figura fondamentale della rivoluzione cubana, hanno rivelato che apparteneva allaplotruppo R1b-L21.
Altri membri famosi dellaplogruppo R1b-L21
- John C. Calhoun (sottoclade M222 > FGC19846): fu il settimo vicepresidente degli Stati Uniti. Viene ricordato soprattutto per la sua strnua difesa della schiavitù. Le sue convinzioni e i suoi moniti nel 1860-61 influenzarono pesantemente la secessione del Sud dallUnione. Il suo aplogruppo è stato determinato analizzando dei discendenti grazie al Calhoun Surname Project.
- Henry Louis Gates (sottoclade M222): un critico letterario americano, educatore, studioso, scrittore ed editore. Fu il primo afroamericano a ricevere la Andrew W. Mellon Foundation Fellowship. Ha ricevuto numerose lauree ad honorem e premi per le sue lezioni, per la sua ricerca e per lo sviluppo degli studi accademici sulla cultura nera. È il presetnarore della serie televisiva Faces of America e Finding Your Roots in onda sulla PBS, che usano le tradizionali ricerche genealogiche e la genetica per scoprire le storie famigliari di Americani noti, inclusi i loro aplogruppi.
- Bill O'Reilly (sottoclade M222): presentatore televisivo americano, scrittore, gionralista, editorialista e commentatore politico. È il presentatore di un programma di approfondimento politico, il The OReilly Factor in onda su Fox New Channel. Il suo aplogruppo Y è stato rivelato dalla serie televisiva Finding Your Roots.
- Bill Maher (sottoclade M222): comico americano, scrittore, produttore, commentatore politico, attore, critico dei media e presentatore televisivo. È noto come presentatore del talk politico della HBO Real Time with Bill Maher. Il suo aplogruppo Y è stato rivelato dalla serie televisiva Finding Your Roots.
- Rory Bremner (sottoclade M222): imitatore e comico scozzese, noto per il suo impegno nella satira politica e per le sue imitazioni di personaggi famosi britannici.
- Adrian Grenier (sottoclade M222): attore americano, produttore, regista e musicista. È noto soprattutto per aver recitato la parte di Vincent Chase nella serie della HBO Entourage.
Ramo R1b-U152 (italo-celtico)

asandosi sui dati dell'Asburgo Family Project, è molto probabile che laplogruppo della casa d'Asburgo sia R1b-U152 (L2+ branch). Gli Asburgo ebbero origine nelleponimo villaggio della Svizzera settentrionale. Divennero Re dei Romani per la prima volta nel 1273, ottenendo il titolo di duchi dAustria. Dagli Asburgo sono poi arrivati tutti i Sacri Romani Imperatori, tranne uno, dal 1440 fino alla dissoluzione dellimpero nel 1806. La loro dinastia regnò anche sulla Spagna, su Napoli, sulla Toscana, su Parma e su Milano. Per conoscere le linee di DNA mitocondriale di molti membri della famiglia degli Asburgo controllate anche Aplogruppi dei re europei e delle regine.

Anallizzando la linea paterna di Riccardo III dInghilterra per confermare lidentità dei suoi presunti resti (vedi Aplogruppo G2a), i ricercatori hanno scoperto che tre suoi parenti moderni, portatori il cognome Somerset e discendenti dalla Casa di Lancaster appartengono tutti allaplogruppo R1b-U152 (x L2, Z36, Z56, M160, M126 e Z192). Nonostante questo indichi una non-paternità in qualche punto della linea dei Plantageneti, è possibile che molti se non tutti i Duca di Beaufort, e possibilmente molti dei monarchi Plantageneti al di fuori della casa di York, appartengano allaplogruppo R1b-U152.

Il primo Presidente degli Stati Uniti, George Washington (1732-1799) potrebbe aver fatto parte della sottoclade L2 di U152. Questa deduzione è basata sui risultati del Washington DNA Project. I suoi antenati arrivarono dallOxfordshire, e prima ancora dal Lancashire. Washington è un cognome abbastanza raro e la sua distribuzione originaria abbraccia proprio queste due regioni, almeno secondo un censimento del 1881. Tutti i membri del progetto con antenati in queste regioni condividono lo stesso aplotipo, cosa che indicherebbe una antenato paterno comune.

Abraham Lincoln (1809-1865), il sedicesimo Presidente degli Stati Uniti, pare appartenesse a R1b-S20376 (U152>L2>Z142>Z150>S20376). Il Lincoln DNA Project ha analizzato diversi discendenti di Samuel the weaver Lincoln, che fu il nonno del trisavolo di Abraham Lincoln. Condividono tutti lo stesso aplotipo.

Grover Cleveland (1837-1908) il ventiduesimo e ventiquattresimo Presidente degli Stati Uniti potrebbe aver fatto parte del ramo L20, una sottoclade di L2, almeno secondo i numerosi risultati del Cleveland DNA Project.
Altri membri famosi dellaplogruppo R1b-U152
- Jörg Jenatsch (1596-1639) : fu un leader politico svizzero durante la Guerra dei Trentanni, una delle figure principali della problematica storia dei Grisoni del XVII secolo.
- Mario Batali : è un cuoco, scrittore, ristoratore e personalità pubblica americana. È un esperto della storia della cultura della cucina italiana, incluse le sue variazioni regionali. Il suo aplogruppo è stato rivelato in Faces of America.
Ramo R1b-DF27 (celtico)
")
Il compositore, teorico musicale e pittore austriaco Arnold Schönberg (1874-1951) apparteneva all'aplogruppo R1b secondo il suo profilo su Geni. Secondo il progetto Schönberg DNA, il suo lignaggio (nipote di Abraham Schoenberg, nato nel 1812 a Szécsény) rientra sotto il clade BY16148 a valle di DF27.
Ramo da determinare

PIl professor Tomasz Kozłowski ha analizzato il DNA Y del principe Janusz II di Masovia, duca di Czersk, di Varsavia, di Liw, di Zakroczym e di Nur. Fu un diretto discendente della dinastia dei Piast, la prima dinastia storica ad aver comandato in Polonia a partire dal Principe Mieszko I (930-992) fino al 1370 con la morte del Re Casimiro III il Grande. La famiglia Piast ha mantenuto il controllo del ducato di Masovia fino al 1526 e del ducato di Silesia fino al 1675. Il prof. Kozłowski ha annunciato che il principe apparteneva allaplogruppo R1b, e come lui molto probabilmente anche altri membri della linea reale, inclusi tutti i duchi di Masovia (1138-1526), sempre se si escludono eventi di non-paternità.

Bogdanowicza et al. (2009) ha analizzato il DNA Y e mitocondriale dei resti riesumati dellastronomo del Rinascimento Niccolò Copernico. Ha così stabilito che apparteneva allaplogruppo Y R1b-P310 e alla linea mitocondriale H.

Rogaev et al. (2009) hanno analizzato il DNA ritrovato nella presunta tomba dello Zar Nicola II di Russia e dei suoi cinque figli. Questi campioni sono stati comparati con campioni di sangue appartenuti a Nicola II e conservati in archivio, così come a campioni presi da discendenti di entrambe le linee paterne e materne. I risultati confermano in modo inequivocabile che la tomba è effettivamente quella di una delle ultime famiglie reali russe. Nicola II apparteneva allaplogruppo Y R1b e allaplogruppo mitocondriale T2. Di conseguenza, tutti gli imperatori russi della dinastia Romanov a partire da Pietro III (1728-1762) appartenevano a questo aplogruppo. Questa linea paterna discende dalla Casa di Oldenburg, a quale include tutti i re di Danimarca a partire da Cristiano I (che regnò dal 1448) così come diversi re di Norvegia, di Svezia e di Grecia, e gli attuali eredi al trono britannico (i principi William e Carlo).

Il grande naturalista inglese Charles Darwin (1809-1882), il quale propose la teoria scientifica dellevoluzione e il processo di selezione naturale, fu un membro dellaplogruppo R1b, almeno secondo i risultati delle analisi di un suo tris nipote.

Kevin Bacon (nato nel 1958), un attore e musicista americano i cui film più importanti includono il musical Footloose (1984), il controverso thriller JFK (1991), il dramma A Few Good Men (2003), il film documentario Apollo 13 (1995) e Mystic River (2003). Bacon ha vinto un Golden Globe e tre Screen Actors Guild Award, ed è stato nominato per un Primetime Emmy Award. Il Guardian lo ha nominato uno dei migliori attori che non ha mai ricevuto una nomination allOscar.

Lattore americano Robert John Downey Jr. (nato nel 1956) ha rivelato il suo aplogruppo Y durante la serie televisiva della PBS Finding Your Roots. Suo padre ha preso il cognome Downey, ma prima di emigrare in America i suoi antenati paterni erano chiamati Elias o Eliasovitz. Ha ricevuto una nomination allOscar per Miglior Attore, ha vinto il BAFTA per il Miglior Attore grazie al suo ruolo di protagonista nel film Chaplin (1992) e ha vinto un Golden Globe per il suo ruolo nella serie TV Ally McBeal. È noto anche per aver interpretato il supereroe della Marvel Iron Man.
Altri membri famosi dellaplogruppo R1b
- Harry Connick Jr. (born 1967) : (nato nel 1967): è un cantante, musicista e attore americano. Ha venduto oltre 28 milioni di album in tutto il mondo. Connick è tra i 60 best-selling artisti maschili degli Stati Uniti, almeno secondo la Recording Industry Association of America.
- Charlie Rose (born 1942) : (nato nel 1942): è un presentatore televisivo americano e un giornalista. Dal 1991 ha presentato Charlie Rose, uno show distribuito dalla PBS dal 1993 dove intervista diversi personaggi.
A seguire
Ask your questions and discuss about haplogroups on the Forum
|



 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")





 in Europa")


 in Europa")
 in Europa")
 in Europa")
















")







