You oppose the theory of steppe migration, but the combined archaeological and ancient DNA evidence strongly supports large scale migrations ultimately derived from Pontic–Caspian steppe populations during the Late Neolithic and Early Bronze Age. What it does not support is a simplistic model in which every later steppe derived population was merely a direct continuation of the classic Yamnaya horizon.
The Yamnaya cultural horizon (~3300–2600 BCE) was heavily associated with R1b-Z2103 paternal lineage, yet the major later expansions into different parts of Europe were carried by populations that had already diverged genetically and culturally from the earliest sampled Yamnaya groups. Corded Ware populations were overwhelmingly associated with R1a-M417, while Bell Beaker groups in western and central Europe became dominated by R1b-P312 downstream of R1b-L51. Early sampled Yamnaya males did not carry either R1a-M417 or R1b-P312. This means the later European expansions were not literal demographic continuations of specific sampled Yamnaya clans.
However, this does not invalidate the broader steppe migration model. Genome wide ancient DNA consistently shows that Corded Ware and Bell Beaker populations derived a very large proportion of their ancestry from populations genetically closest to Yamnaya steppe pastoralists. This is why scholars continue to describe these processes as “Yamnaya derived” or “steppe derived” migrations.
After ~3000 BCE, Europe experienced one of the largest ancestry turnovers ever documented in ancient DNA studies. Corded Ware individuals often derive roughly 70–75% of their ancestry from steppe related populations. Earlier Neolithic farmer associated Y-chromosome lineages declined sharply while new paternal lineages spread across enormous territories. Archaeology independently supports these demographic shifts through shared burial customs, kurgan traditions, pastoral economies, wagon technology and horse use. Linguistically, the spread of many Indo-European languages is also broadly consistent with these movements.
The evidence does not suggest that Yamnaya themselves directly conquered all of Europe. Rather, it supports major migrations carried out by populations descended from or closely related to steppe pastoralists ancestral to later Corded Ware and Bell Beaker groups. That distinction matters because Corded Ware likely formed from earlier steppe derived populations related to, but not identical with, classic Yamnaya. Bell Beaker was also culturally heterogeneous and involved substantial local admixture. Different parts of Europe experienced different demographic processes.
This is also why Y-haplogroups alone cannot define an entire population or migration event. Populations can preserve strong autosomal continuity with earlier steppe ancestors even while dominant paternal lineages shift through founder effects, elite dominance, clan expansion, or breeding bottlenecks. The fact that classic Yamnaya males were not R1a-M417 or R1b-P312 therefore does not disprove the broader steppe migration model. It simply suggests that multiple branches of steppe derived populations expanded in different directions and that currently sampled Yamnaya groups do not represent the full diversity of the wider steppe world ancestral to later Europeans.
The mainstream archaeogenetic position today is that there were major steppe derived migrations into Europe during the 3rd millennium BCE with enormous demographic impact, but Corded Ware and Bell Beaker were not simply identical copies of Yamnaya society. The relationship is best understood as descent from Yamnaya-like or steppe related ancestral populations rather than literal continuity in every genetic, cultural, or paternal lineage detail.
It’s not that I deny steppe migrations per se — what I’m specifically rejecting are Yamnaya migrations as they are commonly portrayed. The problem is that an oversimplified narrative has spread across the internet, and it’s becoming increasingly inaccurate over time.
A story was built around a very specific set of data: the Yamnaya were 70–90% M269>, the Bell Beakers were 70–90% M269>, and they shared roughly 30% similar autosomal ancestry on average. Back in 2018, it was completely coherent and rational to think they were sister populations. That idea became hugely popular and massively boosted DNA test sales, especially in Northern, Central, and Eastern Europe.
Despite the current North/South sampling bias, southern P312> populations (Portugal, Spain, France, Italy) consistently show older average TMRCAs across most clades than northern populations (England, Germany, the Netherlands, Belgium, Austria, Switzerland, Czechia).
The problem is that no P310>, L151>, or P312> has ever been found in the steppes. The closest thing to that lineage is an L51 sample dated to 2800 BC, while L151> itself dates to around 3200 BC — a 400-year gap.
If the western branch really came from that region, it must have migrated before the Yamnaya even existed, sometime around or before 3300 BC.
M269 predates 4500 BC, and the first empirical M269>L23>Z2103 sample dates to around 3800 BC near the North Caucasus. There is also a clear phylogenetic split between the Z2103 clades found in Armenia/Turkey and those from the steppes, meaning we cannot even say with certainty whether M269 originated in the steppes and moved into the Caucasus, or originated in the Caucasus and later spread into the steppes.
So in reality, the most precise statement we can make is that this haplogroup originated somewhere around the Black Sea region — not specifically in the steppes, nor culturally within Yamnaya.
Eight years later, the general summary no longer fits any of its original pillars, and yet the narrative continues rolling forward with the same inertia. The Yamnaya were not responsible for the origin of the Indo-European language, horses, or bronze.
Many people in Northern Europe argue that the earliest L151> samples are found in the Corded Ware Culture of Bohemia, citing Papac et al., but those samples date to around 2800 BC. Some already carry the U106 mutation, and personally I’d say all of them probably do. They are not the direct ancestors of P312*, because they do not share the same STR profiles, and P312* had already existed for at least 200 years by then, even if no equally old aDNA samples have yet been found.
The North can plausibly defend an origin for L51>L151>U106, but not for P312*. In Bell Beaker culture, the elites were overwhelmingly P312>, not U106*. U106 has actually been found very rarely compared to P312 within Bell Beaker contexts.
The archaeological Bell Beaker expansion follows this sequence:
2900 BC Portugal → 2700 BC Spain → 2600 BC France/Italy → 2500 BC Rhine region → 2400 BC British Isles/Hungary.
The oldest P312* samples currently known date to around 2600 BC. We can move radiocarbon dates slightly one way or another, but both major branches are effectively equally ancient because both diverged from P312>ZZ11 roughly 200 years earlier.
~2600 BC Central Europe: P312>ZZ11>U152>L2
~2600 BC Iberia: P312>ZZ11>DF27>ZZ12_1
Their shared male ancestor would be ZZ11 about 200 years earlier. They barely even share IBD segments with each other, so you cannot claim that either branch is older than the other — they are parallel lineages.
They are part of the origin, not the origin itself.
These are the two most successful early Bell Beaker branches, each with more than 30 consolidated immediate lineages.
The founder effect of P312>L21>DF13 is largely irrelevant because it only truly dominated the British Isles and is 100–200 years younger than its older sibling branches. DF13 could indeed be native to the Rhine region or French Brittany.
Are the oldest U152> branches currently found in the North?
No — and they barely reach 5–10% frequencies there.
The oldest U152* branches are found in Italy:
L2, Z36, Z56, PF6585.
Frequencies of at least 15% across the country, peaking above 50% in some northern regions.
Are the oldest DF27> branches currently found in the North?
No — and they also barely reach 5–10%.
The oldest branches are in the Iberian Peninsula:
Z195, ZZ12_1, Z198, Z274, FTT1, FTT3.
Frequencies of at least 40% in almost every region, peaking around 70% among Basques.
It is therefore unlikely that P312* originated in the North, since the clades mentioned above now account for over 70% of all P312> lineages.
Bell Beaker culture emerged in Portugal around 2900 BC.
It is very unlikely that significantly older P312* aDNA samples from 2900–2600 BC will suddenly appear elsewhere, because during that period these lineages were still emerging. Even after Akbari added 15,000 new samples, nothing older than the existing ~2600 BC samples has been found.
So the overall picture is unlikely to change for many years, and the oldest documented P312* samples will probably continue to be EHU002 from Burgos and the U152>L2 samples from Central Europe.
To explain the modern genetic landscape of Iberia, there is no need for Yamnaya invasions, Central European Bronze Age invasions, or Celtic Iron Age invasions.
DF27 is statistically the elder brother of U152 and the oldest clade among all P312>, and this simply cannot be explained through a northern origin model.
Returning to horses: a specimen carrying haplogroup P0* (the only documented example of that lineage outside Iberia) was found near the Nordic region and dated to 3000 BC. In the opposite direction, no horses originating from the North have been found in Iberia.
Then there’s the case of D* haplogroup horses in southwestern Iberia, which diverged from steppe horses around 4000–3500 BC.
What I pointed out in the summary is that humans domesticated horses through multiple independent processes. But selecting animals to create a bulldog (for meat) is not the same as breeding racing greyhounds.
The three major branches of haplogroup D> diverged long before the Sintashta spread their specific lines around 2000 BC. Yet modern horses do not descend directly from those Sintashta lines when examined in detail. So they may have contributed to domestication, but they were neither the beginning nor the end of the process.
The fact that all three branches eventually became 100% DOM2 autosomally through selective breeding cannot be random. It strongly suggests that at least three distinct populations were continuously involved in horse domestication from the beginning.
The problem with that article is that it tries to redefine domestication by mixing bulldogs with racing greyhounds, while ignoring the fact that there are actually two bottlenecks, not one:
the first between 4000–3000 BC, and the second around 500 BC.
The steppe horses of 2000 BC are something intermediate, but they do not explain the entirety of horse domestication, nor are they the starting point. To justify that claim, researchers relied on distinctive genetic markers linked to anxiety and spinal traits, but those lineages were not ultimately the ones that won out. In other words, the horse type we are truly looking for did not really exist before 1000–500 BC.
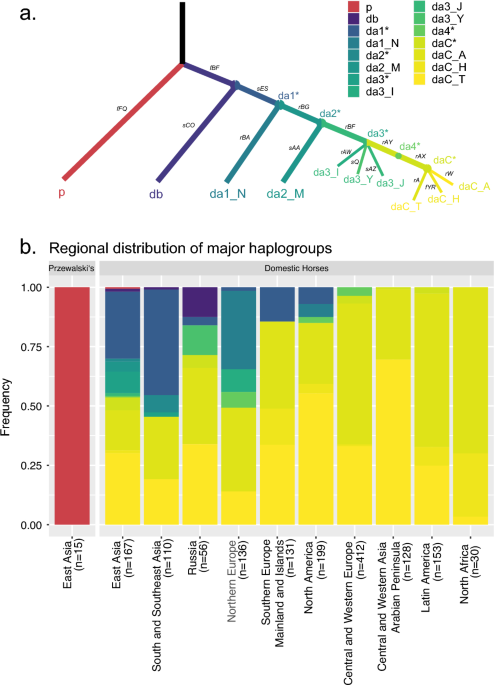
In Iberia we have P0*, D*, DB*, DA*, DA1*, and DAC* — many distinct lineages — yet ultimately only one became dominant. That process took at least 3,000 years for a single line to become globally dominant.
Three thousand years leaves plenty of room for change, and the sampling bias is enormous. The next paper could radically alter the picture with just 20 additional samples. But what can no longer change is the fact that western-origin DAC lineages eventually replaced the eastern ones.
I think it will take several more years before we truly understand which human haplogroups inhabited which regions during each period, and what kinds of horses each group possessed.
M269> appears in every scenario involving horse expansion across different periods, but Z2103> does not.
M269 is not an ordinary haplogroup. It is the macro-haplogroup with the greatest reproductive success in human history within just 6,000 years, potentially having more than 350 million male descendants alive today. That is why it is the most documented and studied haplogroup.
M269>PF7562 and M269>L23>L51>PF7589 also do not appear to be of steppe origin. So far they seem clearly Balkan/Anatolian in origin as well (around the Black Sea region).
The R haplogroups are not solely responsible for Indo-European languages — J2 was also involved. As more data emerges, the evidence increasingly points toward an Anatolian/Caucasian origin rather than a purely steppe one.
So where did P310>L151 really come from?
Probably somewhere in the Balkans/Black Sea region between 3500–3000 BC.
I’m not proposing a radically different origin — only a temporal shift roughly 500 years earlier than currently estimated. But that changes the expansion process dramatically. We would no longer be talking about a massive migration involving many unrelated clans, but rather a founder effect of P312> already established within Atlantic/Mediterranean Europe.
The geographic origin itself does not change, and this also explains the arrival of CHG ancestry in southern Bell Beaker zones. EHG ancestry was already present at around 10–15% among Chalcolithic Iberian I2 groups, and those I2 lineages neither disappeared nor migrated elsewhere (in fact, I’m looking at one right now). P312> lineages simply achieved greater reproductive success after the Bell Beaker period, and that mixture is precisely why modern Iberians remain among the closest populations to the ancient WHG cluster.
The lineages that truly disappeared after the Neolithic were R1B-V88 and H. But the French paper by Buri seems to suggest this had more to do with climate and disease than with the arrival of P312>, and not with any conquest-related massacre.
It is essential to focus strictly on the development of Y-chromosome lineages in both humans and animals, because only the Y chromosome clearly traces the distinct path of each group.
For example:
If CWC populations were “Yamnaya” simply because they cluster autosomally, does that mean the Etruscans were Iberians?
We all know the common factor in both cases is P312>ZZ11, but U152 and DF27 diverged between 2800–2600 BC. What makes them autosomally close is first the Cardial culture and second the Bell Beaker culture.
The Italics do not descend from Iberians, nor Iberians from Italics, yet you can model a modern Balearic individual as 50% Etruscan and an ancient Etruscan as 50% modern Catalan.
The only reason we know they are not recently related is because of the Y chromosome. Without it, people would be speculating wildly with anachronistic interpretations.
There are many people using G25 estimates without understanding how qpAdm actually works, and that is a serious problem.
An even bigger problem is that some people are using qpAdm while presenting models with p-values of 0.9 in aDNA contexts — when achieving 0.9 is usually a sign that something is being overfitted because it is unrealistically precise — while the source Z-scores often do not even exceed 3.
In the end, they are only partially “reading” what they are analyzing, and in most cases doing so anachronistically.
Modeling a single person with ancient samples is relatively easy because of chance: sometimes there simply happen to be ancient populations genetically close to you, and the results look excellent.
The real challenge is explaining entire populations.
If the person doing the modeling does not understand the deep phylogenetic and historical background of each population source, they are basically fooling themselves until statistics eventually show them what they wanted to see from the beginning.
The correct order of importance for investigating the origin of male groups is:
- Y chromosome SNPs
- STR profiles to differentiate unclassified or extinct SNPs
- IBD relationships
- Autosomal modeling with qpAdm, sample by sample
Ordinary people prioritize G25 autosomal modeling instead.
There are many studies with terribly flawed models that are still used because, even if they are not fully accurate, nothing better can currently be achieved due to the lack of intermediate samples. They are still necessary to analyze the available data.
The real problem comes when some people extrapolate that the more “steppe” ancestry you have, the more “Yamnaya” you are — and suddenly they imagine themselves riding a horse with an irresistible urge to conquer Western Europe.
Between P310>L151>P312 there are 500–1000 years that remain unknown, and researchers have tried to fill that gap with the M269>Z2103>KMS67 lineages from the steppes — but the theory simply does not fit together the way they expected.